
Sinh lý học Sinh sản Nam giới
Dịch thuật: Bác sĩ Phú Tiết Niệu
Trục nội tiết và các cơ quan sinh sản nam giới là một hệ thống sinh học được phối hợp nhịp nhàng và quản lý chính xác, đã tiến hóa qua hàng triệu năm. Hệ thống này chịu trách nhiệm hình thành và phát triển đường sinh dục, tiềm năng sinh sản khi bước vào tuổi dậy thì, và duy trì các đặc tính nam giới trưởng thành. Chương này khám phá hiểu biết hiện tại của chúng ta về hệ thống phức tạp này bằng cách định nghĩa giải phẫu và sinh lý học của nó, bao gồm trục nội tiết hạ đồi - tuyến yên - tuyến giáp (HPG) [Ghi chú dịch thuật: văn bản gốc ghi (HPG) và liệt kê tuyến sinh dục (gonadal) ở ngay câu sau, đây có thể là lỗi đánh máy của sách gốc; đúng thuật ngữ chuyên môn HPG là Hạ đồi - Tuyến yên - Tuyến sinh dục], quá trình sinh tinh, sự sản xuất androgen bên trong tinh hoàn, cũng như sự trưởng thành và vận chuyển của tinh trùng trong hệ thống ống dẫn. Ngoài ra, các khái niệm mới về vô sinh do di truyền và biểu sinh (epigenetic), khoa học tế bào gốc, và sinh lý học phóng tinh cũng được giải thích. Thông qua sự mổ xẻ trí tuệ nghiêm túc như vậy, vẻ đẹp và sự tinh tế thực sự của quá trình sinh sản sẽ được trân trọng.
TRỤC HẠ ĐỒI - TUYẾN YÊN - TUYẾN SINH DỤC (HPG)
Trục HPG đóng vai trò quan trọng trong suốt quá trình phát triển và ở độ tuổi trưởng thành đối với bốn quá trình sinh lý: (1) phát triển giới tính kiểu hình trong giai đoạn phôi thai, (2) trưởng thành tình dục ở tuổi dậy thì, (3) chức năng nội tiết của tinh hoàn — sản xuất testosterone, và (4) chức năng ngoại tiết của tinh hoàn — sản xuất tinh trùng.
Các khái niệm nội tiết cơ bản
Có hai loại hormone trung gian điều hòa sự giao tiếp trong trục sinh sản: peptide và steroid. Các hormone peptide là những protein bài tiết nhỏ, hoạt động thông qua các thụ thể trên bề mặt tế bào. Tín hiệu hormone được truyền dẫn bởi một trong số vài con đường truyền tin thứ hai (Hình 64.1). Hầu hết các hormone peptide đều kích hoạt quá trình phosphoryl hóa các protein làm thay đổi chức năng tế bào. Ví dụ về hormone peptide là hormone tạo hoàng thể (LH) và hormone kích thích nang trứng (FSH). Ngược lại, các hormone steroid có nguồn gốc từ cholesterol. Chúng không được lưu trữ trong các hạt bài tiết; do đó, sự bài tiết steroid phản ánh trực tiếp tốc độ sản xuất hormone. Trong huyết tương, các hormone này thường liên kết với các protein vận chuyển, và vì chúng có tính ưa lipid (lipophilic) nên các hormone steroid có thể thấm qua màng tế bào. Sau khi liên kết với các thụ thể nội bào, các steroid được chuyển vị đến các vị trí nhận diện DNA trong nhân và điều hòa quá trình phiên mã của gen đích. Ví dụ về các hormone steroid của trục sinh sản là testosterone và estradiol.
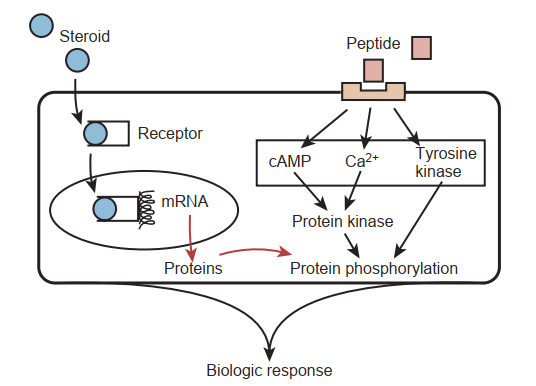
Tín hiệu hormone trong trục HPG được quản lý theo thứ bậc bởi một bộ tạo xung hoạt động tự do nằm trong vùng hạ đồi. Biên độ và tần suất của các đợt bài tiết hormone xảy ra trong trục sinh sản sẽ quyết định khả năng đáp ứng của cơ quan đích hạ lưu. Kiểm soát điều hòa ngược (feedback) là cơ chế chính để quá trình điều hòa hormone diễn ra (Hình 64.2). Với sự kiểm soát điều hòa ngược, một hormone có thể tự điều hòa quá trình tổng hợp và hoạt động của chính nó hoặc của một hormone khác. Trong trục HPG, hoạt động điều hòa ngược âm tính chịu trách nhiệm chính trong việc giảm thiểu các rối loạn và duy trì trạng thái cân bằng nội môi.
Các thành phần của Trục sinh sản
Hạ đồi
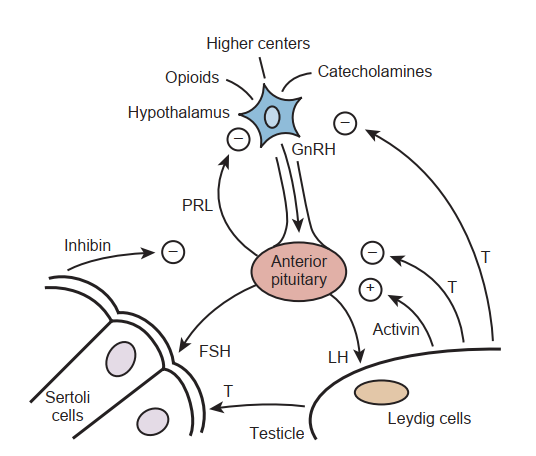
Với vai trò là trung tâm tích hợp của trục HPG, vùng hạ đồi nhận các tín hiệu thần kinh đầu vào từ hạnh nhân (amygdala), đồi thị, cầu não, võng mạc, vỏ não khứu giác và nhiều vùng khác (xem Hình 64.2). Là bộ tạo xung cho sự bài tiết theo chu kỳ của các hormone tuyến yên, vùng hạ đồi được liên kết về mặt giải phẫu với tuyến yên bằng một hệ thống mạch cửa và các con đường thần kinh. Bằng cách tránh tuần hoàn hệ thống, hệ thống mạch cửa cho phép đưa trực tiếp các hormone vùng hạ đồi đến tuyến yên trước.
Hormone vùng hạ đồi quan trọng nhất đối với sinh sản là hormone giải phóng gonadotropin (GnRH), một peptide gồm 10 axit amin được tạo ra trong các thân tế bào thần kinh ở nhân trước thị (preoptic) và nhân cung (arcuate). Hiện tại, chức năng duy nhất được biết đến của GnRH là kích thích sự bài tiết LH và FSH từ tuyến yên trước. GnRH có thời gian bán thải trong huyết tương khoảng 5 đến 7 phút và gần như bị loại bỏ hoàn toàn ngay trong lần đầu tiên đi qua tuyến yên, thông qua cơ chế nội bào hóa thụ thể hoặc bị phân hủy bởi enzyme. Sự bài tiết GnRH là kết quả từ sự tích hợp các yếu tố đầu vào như căng thẳng (stress), tập luyện và chế độ ăn uống từ các trung tâm não cao hơn, các gonadotropin được bài tiết từ tuyến yên và các hormone sinh dục lưu thông trong máu. Các chất được biết đến có khả năng điều hòa sự bài tiết GnRH được liệt kê trong Bảng 64.1. Trong hội chứng Kallmann, đặc trưng bởi tình trạng suy sinh dục do giảm gonadotropin bẩm sinh, các tế bào thần kinh tiền thân GnRH không thể di cư bình thường, dẫn đến sự thiếu hụt bài tiết GnRH của vùng hạ đồi sau đó (Bick et al., 1992; Dode et al., 2003). Những cá thể bị ảnh hưởng sẽ bị chậm dậy thì hoặc vô sinh do không sản xuất được testosterone.
| Tác nhân điều biến GnRH | Loại điều hòa ngược | Ví dụ |
| Opioids | Âm tính | beta-Endorphin |
| Catecholamines | Thay đổi | Dopamine |
| Hormone peptide | Âm tính | FSH, LH |
| Steroid sinh dục | Âm tính | Testosterone |
| Prostaglandins | Dương tính | PGE2 |
| Insulin | Dương tính | Insulin |
| Kisspeptins | Dương tính | Kisspeptin (tuổi dậy thì) |
| Leptins | Dương tính | Leptin |
[BẢNG 64.1 Các chất điều hòa sự bài tiết Hormone giải phóng Gonadotropin (GnRH)]- FSH, Follicle-stimulating hormone; LH, luteinizing hormone; PGE2, prostaglandin E2.
Sự bài tiết GnRH thể hiện một số loại nhịp điệu: nhịp điệu theo mùa, trên thang thời gian tính bằng tháng và đạt đỉnh vào mùa xuân; nhịp điệu sinh học ngày đêm (circadian), dẫn đến nồng độ testosterone cao hơn vào các giờ sáng sớm; và nhịp điệu theo mạch xung (pulsatile), với các đỉnh GnRH xảy ra trung bình mỗi 90 đến 120 phút. Tầm quan trọng của sự bài tiết GnRH theo mạch xung trong chức năng trục HPG bình thường được chứng minh rõ ràng bằng khả năng của các chất đồng vận GnRH ngoại sinh (ví dụ: leuprolide acetate) trong việc làm ngừng sản xuất testosterone ở tinh hoàn bằng cách thay đổi sự tiếp xúc của tuyến yên với GnRH từ dạng chu kỳ sang dạng liên tục.
Tuyến yên trước
Nằm bên trong hố yên bằng xương của hộp sọ, tuyến yên có hai thùy: thùy sau và thùy trước. Thùy sau, hay thùy thần kinh (neurohypophysis), bài tiết hai hormone là oxytocin và vasopressin, và được thúc đẩy bởi các kích thích thần kinh. Ngược lại, tuyến yên trước, hay thùy tuyến (adenohypophysis), được điều hòa bởi các yếu tố truyền qua đường máu và là vị trí tác động của GnRH (xem Hình 64.2). GnRH kích thích sự sản xuất và giải phóng FSH và LH thông qua cơ chế phụ thuộc vào dòng dòng chảy ion canxi. Độ nhạy cảm của các tế bào hướng sinh dục (gonadotrophs) của tuyến yên đối với GnRH thay đổi theo tuổi tác và tình trạng hormone của từng cá thể. LH và FSH là các hormone tuyến yên chính điều hòa chức năng tinh hoàn. Chúng là các glycoprotein được cấu tạo từ hai tiểu đơn vị chuỗi polypeptide, được gọi là $\alpha$ và beta, mỗi chuỗi được mã hóa bởi một gen riêng biệt. Tiểu đơn vị alpha của mỗi hormone là giống hệt nhau và tương tự như tiểu đơn vị $\alpha$ của tất cả các hormone tuyến yên khác; các hoạt tính sinh học và miễn dịch được quyết định bởi tiểu đơn vị $\beta$ duy nhất. Cả hai tiểu đơn vị đều cần thiết cho hoạt động nội tiết. Các phân tử đường liên kết với các tiểu đơn vị peptide này, bao gồm các oligosaccharide có gốc axit sialic, có hàm lượng khác nhau giữa FSH và LH và có khả năng là nguyên nhân dẫn đến sự khác biệt về tốc độ thanh thải của chúng trong huyết tương. Các xung bài tiết của LH có tần suất thay đổi từ 8 đến 16 xung trong 24 giờ và biên độ thay đổi từ 1 đến 3 lần. Các kiểu xung này phản ánh chặt chẽ sự giải phóng GnRH. Androgen và estrogen điều hòa sự bài tiết LH thông qua điều hòa ngược âm tính. Trung bình, các xung FSH xảy ra mỗi 1,5 giờ và biên độ thay đổi khoảng 25%. Sự đáp ứng của FSH đối với GnRH khó đánh giá hơn so với LH vì hai lý do: (1) FSH có biên độ đáp ứng nhỏ hơn và thời gian bán thải trong huyết thanh dài hơn, và (2) các protein sinh dục như inhibin và activin có thể ảnh hưởng đến sự bài tiết FSH và được cho là nguyên nhân dẫn đến tính độc lập bài tiết tương đối của FSH đối với sự bài tiết GnRH.
FSH và LH chỉ được biết đến là có tác dụng tại tuyến sinh dục. Chúng kích hoạt adenylate cyclase, dẫn đến sự gia tăng adenosine monophosphate vòng (cAMP) nội bào. Tại tinh hoàn, LH kích thích quá trình tổng hợp steroid bên trong các tế bào Leydig bằng cách kích ứng sự chuyển đổi cholesterol ở ty thể thành pregnenolone và testosterone. FSH liên kết với các tế bào Sertoli và màng tế bào tinh nguyên bào bên trong tinh hoàn, và là chất kích thích chính cho sự phát triển của ống sinh tinh trong quá trình phát triển. FSH là thiết yếu cho sự khởi đầu của quá trình sinh tinh ở tuổi dậy thì. Ở người trưởng thành, vai trò sinh lý chính của FSH là kích thích quá trình sinh tinh đạt các mức độ bình thường về mặt số lượng (Tapanainen et al., 1997).
Một hormone thứ ba của tuyến yên trước là prolactin cũng có thể ảnh hưởng đến trục HPG và khả năng sinh sản. Prolactin là một protein hình cầu lớn gồm 199 axit amin (23 kD), chịu trách nhiệm tổng hợp sữa trong thai kỳ và thời kỳ cho con bú ở phụ nữ. Không có đột biến nào ở người được tìm thấy trong gen prolactin hoặc thụ thể của nó ở người (Goffin et al., 2002). Vai trò của prolactin ở nam giới kém rõ ràng hơn, nhưng nó có thể làm tăng nồng độ của các thụ thể LH trên tế bào Leydig và duy trì nồng độ testosterone cao bình thường trong tinh hoàn. Nó cũng có thể làm tăng cường tác dụng của androgen đối với sự phát triển và bài tiết của các tuyến sinh dục phụ ở nam giới (Steger et al., 1998; Wennbo et al., 1997). Nồng độ prolactin bình thường có thể quan trọng để duy trì ham muốn tình dục. Mặc dù nồng độ prolactin thấp không nhất thiết là bệnh lý, nhưng tình trạng tăng prolactin máu (hyperprolactinemia) sẽ làm mất tính chất mạch xung của gonadotropin bằng cách can thiệp vào sự giải phóng GnRH theo đợt. Ngoài ra, tuyến yên trước còn chứa các tế bào bài tiết các hormone glycoprotein khác: hormone hướng vỏ thượng thận (ACTH), hormone tăng trưởng (GH), và hormone kích thích tuyến giáp (TSH). Các hormone glycoprotein này cũng có thể có những ảnh hưởng đáng kể đến chức năng sinh sản nam giới.
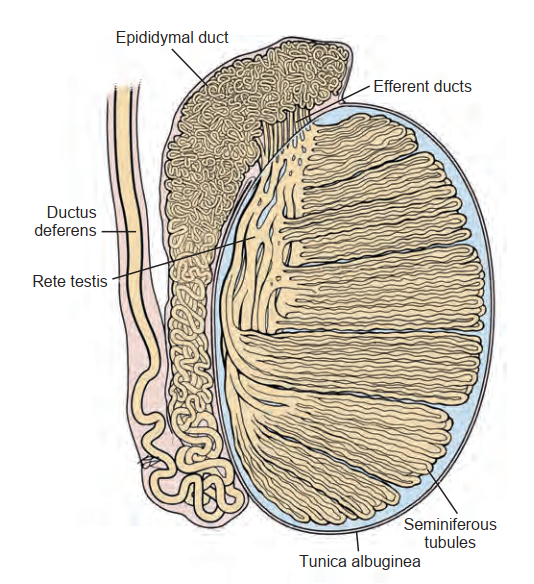
Tinh hoàn
Sự nam tính và khả năng sinh sản bình thường của nam giới đòi hỏi sự hợp tác của cả tinh hoàn ngoại tiết và nội tiết (xem Hình 64.2). Khoang kẽ, cấu tạo chủ yếu từ các tế bào Leydig, chịu trách nhiệm cho quá trình tổng hợp steroid. Các ống sinh tinh sản xuất ra tinh trùng.
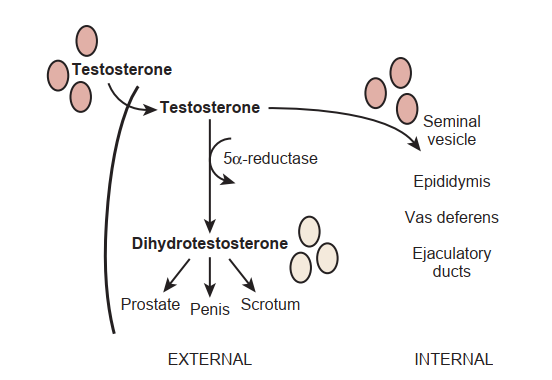
Sự sản xuất testosterone bình thường ở nam giới là khoảng 5 mg/ngày [Ghi chú dịch thuật: văn bản gốc ghi "5 g/day", đây là lỗi in ấn phổ biến trong tài liệu gốc; lượng testosterone sản xuất hàng ngày ở nam giới trưởng thành bình thường là khoảng 5–7 mg/ngày], và sự bài tiết diễn ra theo một cách thức có xung, không đều và giảm dần (nhịp ngày đêm/nyctohemeral). Testosterone được chuyển hóa thành hai chất chuyển hóa hoạt động chính tại mô đích: (1) androgen chính là dihydrotestosterone (DHT) từ hoạt động của enzyme 5-alpha-reductase, và (2) estrogen là estradiol thông qua hoạt động của các enzyme aromatase. DHT là một androgen mạnh hơn nhiều so với testosterone. Ở hầu hết các mô đích ngoại vi, sự khử testosterone thành DHT là bắt buộc đối với hoạt động của androgen, nhưng ở tinh hoàn và cơ xương, sự chuyển đổi thành DHT không phải là điều kiện thiết yếu cho hoạt động của hormone này.
Vị trí tác động chính của FSH là trên các tế bào Sertoli nằm trong các ống sinh tinh. Để đáp ứng với FSH, các tế bào Sertoli sản xuất ra protein liên kết androgen (ABP), transferrin, lactate, ceruloplasmin, clusterin, chất kích hoạt plasminogen, các prostaglandin, và các yếu tố tăng trưởng. Thông qua các yếu tố gián tiếp bởi FSH này, sự phát triển của ống sinh tinh được kích thích trong quá trình phát triển, và sự sản xuất tinh trùng được khởi động trong tuổi dậy thì. Các nghiên cứu bất hoạt gen (knockout) FSH ở chuột gợi ý rằng FSH không phải là yếu tố bắt buộc đối với quá trình sinh tinh; trên thực tế, những con chuột bị ảnh hưởng vẫn có thể có khả năng sinh sản (Levallet et al., 1999).
Tinh hoàn cũng sản xuất các hormone protein là inhibin và activin (Itman et al., 2006). Inhibin là một protein có trọng lượng 32-kD do các tế bào Sertoli tạo ra, có tác dụng ức chế sự giải phóng FSH từ tuyến yên. Bên trong tinh hoàn, sự sản xuất inhibin được kích thích bởi FSH và hoạt động theo cơ chế điều hòa ngược âm tính tại tuyến yên hoặc vùng hạ đồi. Activin, một protein tinh hoàn có sự tương đồng cấu trúc chặt chẽ với yếu tố tăng trưởng chuyển đổi beta (TGF-beta), có tác dụng kích thích lên sự bài tiết FSH. Các thụ thể activin được tìm thấy trong một loạt các mô ngoài tuyến sinh dục, gợi ý rằng hormone này có thể có vai trò là yếu tố tăng trưởng hoặc vai trò điều hòa trong cơ thể.
Sự ức chế điều hòa ngược âm tính đối với sự giải phóng GnRH bởi testosterone xảy ra thông qua các thụ thể androgen (ARs) ở các tế bào thần kinh vùng hạ đồi và trong tuyến yên. Trong các nghiên cứu về đột biến di truyền, rõ ràng là cả testosterone và estrogen đều tham gia vào quá trình điều hòa ngược âm tính (Shupnik và Schreihofer, 1997). Sự điều hòa ngược âm tính của steroid chủ yếu là do sự liên kết của AR với testosterone, với một phần đóng góp nhỏ hơn từ sự liên kết của estradiol. Sự điều hòa ngược của testosterone xảy ra chủ yếu ở vùng hạ đồi, trong khi sự điều hòa ngược của estrogen xảy ra chủ yếu ở tuyến yên (Santen, 1975). Người ta cũng nhận thấy rằng mặc dù testosterone là chất điều hòa chính cho sự bài tiết LH, estradiol (cùng với inhibin từ tế bào Sertoli) là chất điều hòa chiếm ưu thế đối với sự bài tiết FSH (Hayes et al., 2001).
Sự phát triển của Trục Hạ đồi - Tuyến yên - Tuyến sinh dục
Sự xác định giới tính ở người là do di truyền. Một gen quan trọng để xác định giới tính là SRY (gen vùng xác định giới tính Y) nằm trên nhánh ngắn của nhiễm sắc thể Y. Sản phẩm của gen SRY là một protein có trình tự hộp nhóm di động cao (HMG), một mô-típ liên kết DNA được bảo tồn cao có khả năng làm cong DNA. Hiệu ứng bẻ cong DNA này làm thay đổi sự biểu hiện gen, dẫn đến việc hình thành tinh hoàn và tiếp theo là kiểu hình nam giới. Tuy nhiên, gen SRY không hoạt động độc lập để xác định giới tính con người. DAX1, một gen thụ thể hormone trong nhân, có thể làm thay đổi hoạt động của SRY trong quá trình phát triển bằng cách đàn áp các gen hạ lưu của SRY vốn bình thường sẽ kích ứng sự biệt hóa tinh hoàn. Một gen thứ hai là WNT4, phần lớn giới hạn ở buồng trứng trưởng thành, cũng có thể hoạt động như một gen "chống tinh hoàn". Việc phát hiện ra các gen này đã làm thay đổi đáng kể các lý thuyết về xác định giới tính. Trước đây, kiểu gen nữ được coi là con đường phát triển mặc định ("default"), không có SRY. Giờ đây, rõ ràng là các gen như WNT4 và DAX1 có thể chủ động kích ứng sự phát triển tuyến sinh dục nữ, ngay cả khi có sự hiện diện của SRY (DiNapoli và Capel, 2008).
Một khi giới tính tuyến sinh dục được xác định, các tế bào Leydig sẽ sản xuất testosterone, chất kích ứng sự phát triển của cơ quan sinh dục trong (Hình 64.3). Tế bào Leydig cũng tổng hợp yếu tố tăng trưởng giống insulin-3 để thúc đẩy quá trình di chuyển xuyên ổ bụng của tinh hoàn vào trong bìu. DHT nam tính hóa mầm cơ quan sinh dục để hình thành cơ quan sinh dục ngoài (xem Hình 64.3).
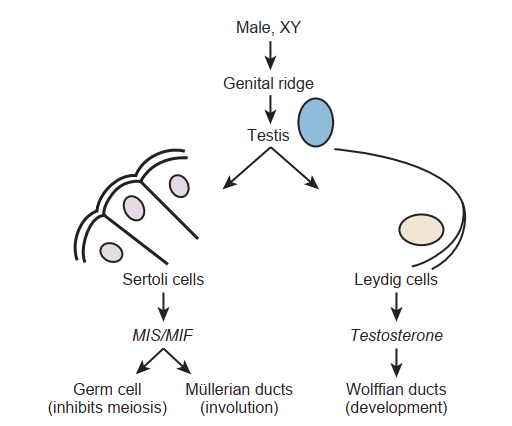
Ngoài ra, các tế bào Sertoli bên trong tinh hoàn đang phát triển tổng hợp chất ức chế müllerian (MIS, hoặc hormone antimüllerian [AMH]), chất này ngăn không cho ống müllerian phát triển thành tử cung và vòi trứng, đồng thời giữ cho các tế bào mầm giai đoạn sớm ở trạng thái nghỉ trong tinh hoàn (Hình 64.4). Nói chung, những khiếm khuyết trong các con đường phát triển này sẽ dẫn đến dị tật bẩm sinh hoặc các rối loạn lưỡng tính.
Các mối quan hệ điều hòa ngược hormone trong trục HPG được thiết lập trong thời kỳ mang thai. Sự biểu hiện của protein kisspeptin chịu trách nhiệm một phần trong việc kích hoạt các tế bào thần kinh GnRH và khởi phát sự giải phóng GnRH. Ngoài ra, SF-1, một thụ thể nhân mồ côi (orphan nuclear receptor), được tiết ra bởi các tế bào Sertoli đang phát triển và đóng góp vào sự phát triển của trục HPG (Val et al., 2003). Sau khi các steroid từ nhau thai rút đi lúc sinh, có một giai đoạn bài tiết gonadotropin cao ở trẻ sơ sinh. Sau đó, khi độ nhạy của trục đối với gonadotropin tăng lên, sự bài tiết FSH và LH giảm xuống các mức thấp đặc trưng của thời thơ ấu. Tuổi dậy thì bắt đầu với việc phát xung GnRH, dẫn đến việc các gonadotropin tăng lên mức của người trưởng thành và sau đó làm tăng các hormone sinh dục.
Khả năng tạo ra các xung GnRH của vùng hạ đồi xuất hiện vào tuổi dậy thì, thường bắt đầu vào khoảng năm thứ 12 ở nam giới. Tuổi dậy thì bắt đầu ở các mức độ tới hạn về tăng trưởng, cân nặng và dinh dưỡng của các bé trai và bé gái, và có khả năng được khởi động bởi kisspeptin, melatonin và leptin (Clement et al., 1998). Hormone leptin từ tế bào mỡ là tín hiệu điều hòa của cơ thể quản lý kích thước của lượng mỡ dự trữ, và ngày càng có nhiều bằng chứng cho thấy leptin điều hòa hoạt động của hạ đồi và tuyến yên (Caprio et al., 1999; Kiess et al., 1999; Quinton et al., 1999).
Sự lão hóa và Trục Hạ đồi - Tuyến yên - Tuyến sinh dục
Sự suy giảm dần trong sản xuất testosterone và tinh trùng xảy ra theo tuổi tác, đến mức nam giới ở thập kỷ thứ bảy có nồng độ testosterone huyết tương trung bình thấp hơn 35% so với nam giới trẻ tuổi (Vermeulen et al., 1995). Hệ quả của điều này là một hiện tượng được gọi bằng nhiều tên khác nhau như mãn kinh nam (male menopause), thời kỳ mãn dục nam (male climacteric), tắt dục nam (andropause), hoặc tên gọi phù hợp hơn là suy giảm androgen một phần ở nam giới lớn tuổi (PADAM). Những thay đổi đối với biểu mô sinh tinh theo tuổi tác bao gồm sự giảm thể tích và chiều dài của ống sinh tinh. Sự sụt giảm sản xuất tinh trùng liên quan đến tuổi tác ở những tinh hoàn già hơn dường như xuất phát từ việc giảm sự tăng sinh tế bào mầm hơn là do tăng sự thoái hóa tế bào. Tương ứng, nồng độ FSH cũng tăng theo tuổi tác, với các giá trị trung bình cao gấp ba lần ở nam giới lớn tuổi so với nam giới trẻ tuổi. Nguyên nhân của sự suy giảm chức năng trục HPG liên quan đến tuổi tác là do đa yếu tố. Sự sản xuất testosterone bị giảm do có ít tế bào Leydig hơn và có nhiều protein liên kết testosterone hơn. Sự thay đổi theo nhịp ngày đêm của bài tiết testosterone cũng bị mất đi ở nam giới lớn tuổi. Theo tuổi tác, cũng có bằng chứng về phản ứng điều hòa ngược của trục HPG bị cùn mòn đối với tình trạng testosterone thấp (bất kể mức độ gonadotropin nhìn chung là cao) và đối với sự kích thích của GnRH. Cuối cùng, sự giải phóng GnRH theo mạch xung bình thường được thay thế bằng các xung không đều, vốn kém hiệu quả hơn trong việc kích thích giải phóng gonadotropin (Mulligan et al., 1997). Sự kết hợp của các hiệu ứng này có khả năng là nguyên nhân gây suy giảm chức năng trục HPG theo tuổi tác.
TINH HOÀN
Cấu trúc đại thể
Tinh hoàn là một cơ quan hình bầu dục, màu trắng, bình thường có thể tích từ 15 đến 25 mL (Prader, 1966) và có chiều dài từ 4,5 đến 5,1 cm (Tishler, 1971; Winter và Faiman, 1972). Lớp màng trắng (tunica albuginea) có các tế bào cơ trơn chạy qua mô collagen (Langford và Heller, 1973). Các tế bào cơ trơn có thể mang lại khả năng co bóp cho bao xơ tinh hoàn (Rikmaru và Shirai, 1972), có thể ảnh hưởng đến dòng máu đi vào tinh hoàn (Schweitzer, 1929), và thúc đẩy dòng chảy của dịch ống sinh tinh ra khỏi tinh hoàn (Davis và Horowitz, 1978).
Nhu mô tinh hoàn được chia thành các ngăn bởi các vách xơ (septa). Mỗi vách xơ chia các ống sinh tinh thành các thùy, mỗi thùy chứa một động mạch ly tâm. Từng ống sinh tinh riêng lẻ là nơi trú ngụ của các tế bào mầm đang phát triển. Mô kẽ được cấu tạo từ các tế bào Leydig, tế bào mast, đại thực bào, dây thần kinh, mạch máu và mạch bạch huyết. Ở người, mô kẽ chiếm 20% đến 30% tổng thể tích tinh hoàn (Setchell và Brooks, 1988). Mối quan hệ giữa các ống sinh tinh và giải phẫu mô kẽ được thể hiện trong Hình 64.5.
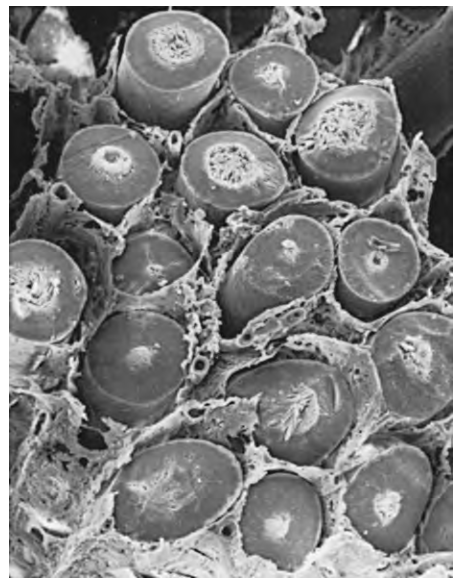
Các ống sinh tinh dài, cuộn xoắn cao và tạo thành các vòng lặp. Cả hai đầu đều tận cùng tại lưới tinh hoàn (rete testis). Tổng chiều dài của 600 đến 1200 ống sinh tinh trong tinh hoàn người được ước tính là 250 m (Lennox và Ahmad, 1970; Hình 64.6). "Trung tâm" của tinh hoàn, còn được gọi là lưới tinh hoàn, hợp lại để hình thành 6 đến 12 ống xuất (ductuli efferentes) có nhiệm vụ mang dịch tinh hoàn và tinh trùng vào đầu mào tinh (caput epididymis) (xem Hình 64.6).
CÁC ĐIỂM CHÍNH: TRỤC HẠ ĐỒI - TUYẾN YÊN - TUYẾN SINH DỤC
- Sự sản xuất testosterone và tinh trùng bình thường phụ thuộc vào sự bài tiết theo mạch xung của GnRH vùng hạ đồi cùng với LH và FSH từ tuyến yên trước.
- Sự điều hòa các hormone trục HPG xảy ra chủ yếu thông qua cơ chế điều hòa ngược âm tính.
- Sự xác định đặc tính nam giới bắt nguồn từ gen SRY trên nhiễm sắc thể Y. Tuy nhiên, các gen phát triển như WNT4 và DAX1 được coi là các gen chống tinh hoàn và có thể chủ động kích ứng sự phát triển tuyến sinh dục nữ.
- Những thay đổi đối với trục HPG theo tuổi của người cha bao gồm nồng độ testosterone thấp hơn, phản ứng điều hòa ngược của trục bị cùn mòn, và tính chất mạch xung hormone không đều.
Nguồn cấp động mạch cho tinh hoàn và mào tinh hoàn bắt nguồn từ ba nguồn: động mạch tinh hoàn trong (internal spermatic artery), động mạch ống dẫn tinh (deferential/vasal artery), và động mạch tinh hoàn ngoài (external spermatic / cremasteric artery) (Harrison và Barclay, 1948). Động mạch tinh hoàn trong xuất phát từ động mạch chủ bụng và liên kết chặt chẽ với đám rối tĩnh mạch hình dây leo (pampiniform plexus). Sự sắp xếp mạch máu trong đám rối hình dây leo, với động mạch và tĩnh mạch có dòng chảy ngược chiều nhau, tạo điều kiện thuận lợi cho sự trao đổi nhiệt và các phân tử nhỏ. Ví dụ, testosterone khuếch tán thụ động từ tĩnh mạch sang động mạch theo cách thức bị giới hạn bởi nồng độ (Bayard et al., 1975). Sự trao đổi nhiệt ngược dòng này cung cấp máu động mạch cho tinh hoàn có nhiệt độ thấp hơn từ 2°C đến 4°C so với nhiệt độ trực tràng ở nam giới bình thường (Agger, 1971). Sự mất đi sự chênh lệch nhiệt độ này có liên quan đến tình trạng rối loạn chức năng tinh hoàn ở nam giới bị giãn tĩnh mạch thừng tinh (Goldstein và Eid, 1989) và tinh hoàn ẩn (cryptorchidism) (Marshall và Edler, 1982). Vì thừng tinh thường được phẫu thuật bóc tách trong quá trình sửa chữa giãn tĩnh mạch thừng tinh, điều liên quan về mặt ngoại khoa cần biết là có một động mạch duy nhất được quan sát thấy trong 50% các thừng tinh, với hai động mạch trong 30% và ba động mạch trong 20% các trường hợp (Beck et al., 1992).
Phía dưới đám rối hình dây leo của bìu và gần trung thất tinh hoàn, động mạch tinh hoàn cuộn xoắn cao và phân nhánh trước khi đi vào tinh hoàn. Các mối nối thông rộng rãi, đặc biệt là giữa động mạch tinh hoàn trong và động mạch ống dẫn tinh, cho phép duy trì khả năng sống của tinh hoàn ngay cả sau khi thắt động mạch tinh hoàn trong (Hình 64.7).
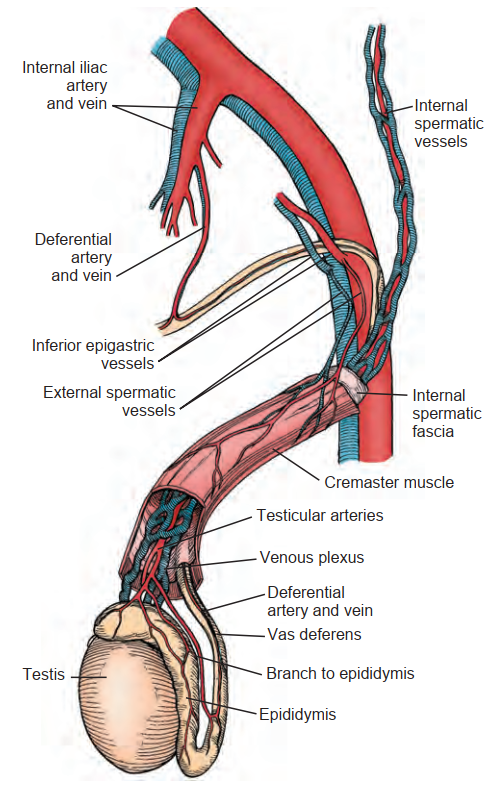
Từ các nghiên cứu chụp mạch, có một động mạch duy nhất đi vào tinh hoàn trong 56% trường hợp; hai nhánh trong 31% trường hợp, và ba hoặc nhiều nhánh hơn ở 13% tinh hoàn (Kormano và Suoranta, 1971). Ở nam giới chỉ có một động mạch tinh hoàn duy nhất, sự gián đoạn của nó có thể dẫn đến teo tinh hoàn (Silber, 1979). Các động mạch tinh hoàn xuyên qua màng trắng và đi xuống dưới dọc theo mặt sau của tinh hoàn bên trong nhu mô. Các động mạch phân nhánh đi về phía trước trên nhu mô tinh hoàn. Các nhánh động mạch tinh hoàn lớn cũng đi qua cực dưới của tinh hoàn, đi về phía trước và phân nhánh trên bề mặt tinh hoàn. Vị trí của các mạch máu này có tầm quan trọng trên lâm sàng, bởi vì chúng có thể bị tổn thương trong quá trình phẫu thuật hạ tinh hoàn (orchiopexy), sinh thiết tinh hoàn, hoặc các thủ thuật thu lấy tinh trùng (Jarow, 1991; Schlegel và Su, 1997). Vùng giữa của tinh hoàn có tương đối ít mạch máu hơn so với các vùng cực trên hoặc cực dưới. Các động mạch riêng lẻ đến các ống sinh tinh, được gọi là các động mạch ly tâm, chạy bên trong các vách xơ. Các nhánh động mạch ly tâm tạo ra các tiểu động mạch cung cấp cho từng mao mạch gian ống (intertubular) và quanh ống (peritubular). Các mao mạch gian ống nằm bên trong các cột mô kẽ, trong khi các mao mạch hình bậc thang chạy gần ống sinh tinh được gọi là các mao mạch quanh ống. Thông qua phức hợp mạch máu này, tinh hoàn người được cung cấp 9 mL máu trên 100 g mô trong mỗi phút (Pettersson et al., 1973).
Các tĩnh mạch bên trong tinh hoàn có điểm bất thường là chúng không chạy cùng với các động mạch tinh hoàn tương ứng. Các tĩnh mạch nhu mô nhỏ đổ vào các tĩnh mạch trên bề mặt tinh hoàn hoặc vào một nhóm tĩnh mạch gần trung thất tinh hoàn chạy dọc theo lưới tinh hoàn (Setchell và Brooks, 1988). Hai nhóm tĩnh mạch này kết hợp với nhau cùng với các tĩnh mạch ống dẫn tinh để tạo thành đám rối hình dây leo khi chúng đi lên vào trong bìu. Các tĩnh mạch đám rối hình dây leo có thành mỏng, điều này có khả năng góp phần vào sự khuếch tán hiệu quả của testosterone và nhiệt với động mạch tinh hoàn nằm liên kết chặt chẽ.
Tinh hoàn không có sự phân bố thần kinh thể xác (somatic innervation) nào được biết đến. Nó nhận sự phân bố thần kinh tự chủ chủ yếu từ các dây thần kinh liên mạc treo và đám rối thận (Mitchell, 1935). Các dây thần kinh này chạy dọc theo động mạch tinh hoàn để đi vào tinh hoàn. Người ta nhận thấy rằng sự phân bố thần kinh giao cảm (adrenergic) của tinh hoàn bị giới hạn chủ yếu ở các mạch máu nhỏ cung cấp cho các cụm tế bào Leydig, có thể điều hòa quá trình tổng hợp steroid của tế bào Leydig (Baumgarten et al., 1968; Turnbull và Rivier, 1997). Người ta cho rằng trương lực mạch máu ở tinh hoàn được điều hòa ở nhiều cấp độ (Linzell và Setchell, 1969), bao gồm sự tự điều hòa của các động mạch bao xơ (Davis et al., 1990), và sự thay đổi theo vùng dựa trên nhu cầu chuyển hóa tại chỗ và được chi phối bởi các peptide như peptide lợi niệu natri tâm nhĩ (Collin et al., 1997) cùng với sự vận chuyển hỗ trợ của các phân tử như LH qua nội mạc mạch máu (Milgrom et al., 1997). Thật vậy, những quan sát này gợi ý một chức năng chuyên biệt cao của hệ vi tuần hoàn tinh hoàn (xem bài tổng quan của Desjardins [1989]).
Có thể quan sát thấy các mạch bạch huyết nổi rõ bên trong thừng tinh (Hundeiker, 1971). Sự tắc nghẽn của các ống này dẫn đến sự giãn nở của mô kẽ tinh hoàn chứ không phải các ống sinh tinh, gợi ý rằng khoảng kẽ được dẫn lưu bởi các mạch bạch huyết. Tắc nghẽn bạch huyết cũng có thể dẫn đến hình thành tràn dịch màng tinh hoàn (hydrocele), một biến chứng đã được biết đến của các phẫu thuật cắt bỏ giãn tĩnh mạch thừng tinh và phẫu thuật khâu thoát vị bẹn.
Dịch trong ống chứa tinh trùng bao quanh các tế bào Sertoli chảy từ các ống sinh tinh vào lưới tinh hoàn và sau đó vào đầu mào tinh. Dịch này, có áp suất thẩm thấu tương đương (isosmotic) với huyết tương, được cho là có nguồn gốc chủ yếu từ ống sinh tinh (Setchell và Brooks, 1988). Sự tái hấp thu dịch này bên trong lưới tinh hoàn và các ống xuất được điều hòa bởi các estrogen (Lee et al., 2000). Thành phần dịch ống khác biệt rõ rệt so với huyết tương hoặc dịch bạch huyết, gợi ý rằng các chất không thể khuếch tán tự do vào và ra khỏi các ống (Setchell và Waites, 1975). Điều này dẫn đến khái niệm về một "hàng rào máu - tinh hoàn" sẽ được thảo luận ở phần sau.
Cấu trúc tế bào tinh hoàn
Mô kẽ
Tế bào Leydig. Mô kẽ tinh hoàn chứa các mạch máu, mạch bạch huyết, nguyên bào sợi, đại thực bào, tế bào mast và tế bào Leydig (Hình 64.8). Các tế bào Leydig chịu trách nhiệm cho phần lớn sự sản xuất steroid của tinh hoàn. Tế bào Leydig biệt hóa từ các tế bào tiền thân trung mô vào tuần thứ bảy của thai kỳ. Sự kích hoạt quá trình tổng hợp steroid của tế bào Leydig tương quan với sự bắt đầu biệt hóa phụ thuộc vào androgen của hệ sinh sản nam. Tế bào Leydig biệt hóa từ các tế bào gốc tiền thân dưới ảnh hưởng của LH và gonadotropin màng đệm người (hCG) có nguồn gốc từ nhau thai, và từ tác động của các yếu tố cận tiết (paracrine) tại chỗ như yếu tố tăng trưởng giống insulin-1 (IGF-1) (Huhtaniemi và Pelliniemi, 1992; Le Roy et al., 1999; Teerds và Dorrington, 1993). Đến 3 tháng sau khi sinh, một làn sóng biệt hóa tế bào Leydig thứ hai xảy ra để đáp ứng với sự sản xuất LH của tuyến yên, làm tăng tạm thời nồng độ testosterone. Androgen được sản xuất trong cuộc sống sơ sinh sớm của trẻ nam được cho là để in vết hormone (hormonally imprint) lên vùng hạ đồi, gan và tuyến tiền liệt để chúng phản ứng phù hợp với sự kích thích của androgen sau này trong cuộc sống. Sau khi tái kích hoạt trục HPG ở tuổi dậy thì, phân tích lập thể đã tiết lộ rằng một tinh hoàn duy nhất từ một người trẻ trưởng thành chứa khoảng 700 triệu tế bào Leydig (Kaler và Neaves, 1978).
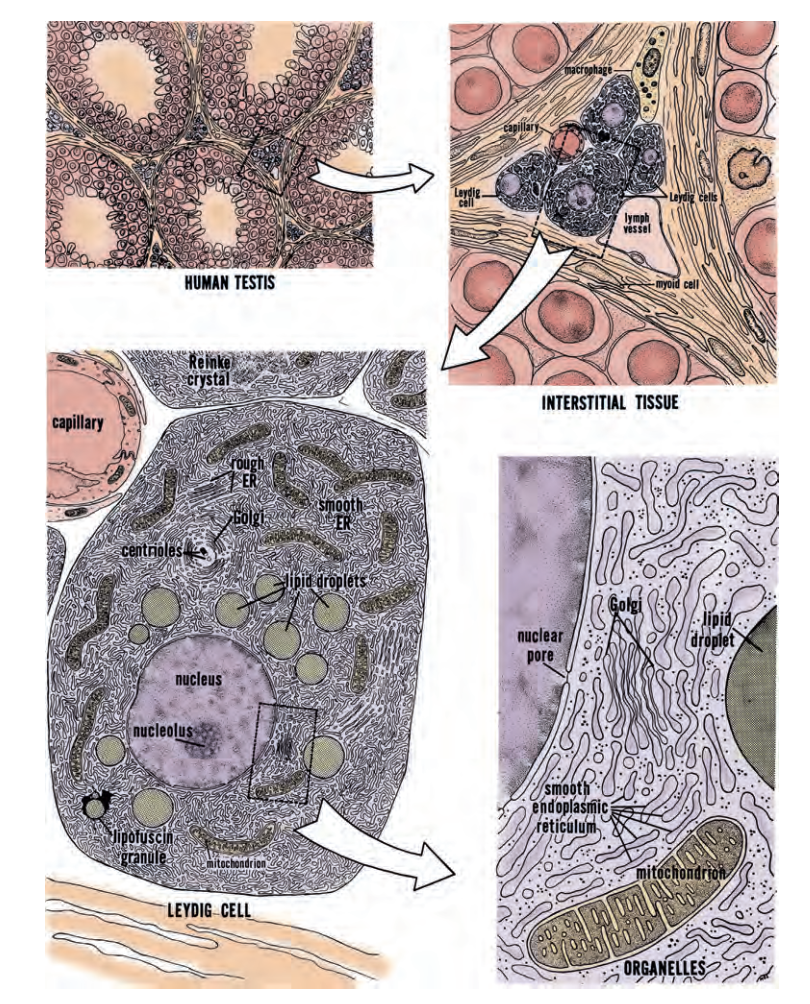
Testosterone. Testosterone, được tổng hợp từ cholesterol, là steroid chính được sản xuất bởi tinh hoàn (Lipsett, 1974). Nhiều steroid C18, C19 và C21 khác cũng được sản xuất (Ewing và Brown, 1977; Lipsett, 1974). Cholesterol được vận chuyển vào trong ty thể của tế bào Leydig, nơi enzyme cắt chuỗi bên cholesterol chuyển đổi nó thành pregnenolone. Việc duy trì các kho dự trữ cholesterol là một phần chức năng bình thường của tế bào Leydig; sự kích thích của LH gợi ra sự huy động cholesterol thông qua hoạt động của enzyme cholesterol esterase. Pregnenolone được vận chuyển ra khỏi màng ty thể vào trong lưới nội chất trơn, nơi nó được chuyển đổi thành testosterone. Testosterone khuếch tán qua màng tế bào và bị giữ lại bên trong dịch ngoại bào và huyết tương bởi các protein liên kết steroid.
Sự vận chuyển cholesterol đến màng trong của ty thể được điều hòa bởi hai protein vận chuyển: protein điều hòa cấp tính tổng hợp steroid (StAR) và thụ thể benzodiazepine ngoại vi (PBR). Sự liên kết của LH gợi ra sự tổng hợp StAR trong tế bào Leydig, protein này sau đó xuyên qua màng ngoài ty thể.
Kiểm soát tổng hợp Testosterone. Sự kiểm soát quá trình tổng hợp steroid của tế bào Leydig rất phức tạp và liên quan đến các yếu tố tuyến yên và ngoài tuyến yên (Payne và Youngblood, 1995). Chất điều hòa quan trọng nhất của sự sản xuất testosterone là LH. Sau khi liên kết với LH, thông qua chất truyền tin thứ hai cAMP, các tế bào Leydig khởi động sự vận chuyển cholesterol vào ty thể. Các peptide tuyến yên khác ngoài LH (ví dụ: FSH và prolactin) làm thay đổi phản ứng đối với LH (Ewing, 1983). Các yếu tố ngoài tuyến yên khác có khả năng làm thay đổi sự sản xuất steroid của tế bào Leydig bao gồm GnRH (Sharpe, 1984); inhibin và activin (Bardin et al., 1989); yếu tố tăng trưởng biểu bì (EGF), IGF-1, và TGF-beta (Ascoli và Segaloff, 1989; Saez et al., 1991); các prostaglandin (Eik-Nes, 1975); và sự kích thích giao cảm (Eik-Nes, 1975). Hơn nữa, sự ức chế trực tiếp quá trình tổng hợp steroid của tế bào Leydig có thể tạo điều kiện thuận lợi cho sự vận chuyển cholesterol (Stocco, 2000). PBR tạo thành một kênh cho cholesterol trong màng ty thể (Culty et al., 1999), nhưng chưa rõ liệu PBR có tương tác chức năng với StAR hay không (West et al., 2001).
Bốn enzyme chính tham gia vào quá trình sinh tổng hợp testosterone từ pregnenolone là enzyme cắt chuỗi bên cholesterol, 3beta-hydroxysteroid dehydrogenase, cytochrome P450 17alpha-hydroxylase/C17-20-lyase, và 17beta-hydroxysteroid dehydrogenase. Động học enzyme, vị trí trên nhiễm sắc thể và di truyền học phân tử của các enzyme này đã được mô tả rõ ràng (Payne và Hales, 2004). Các đột biến trong các gen mã hóa các enzyme này đã được mô tả, và các rối loạn sinh tổng hợp androgen dẫn đến là một nguyên nhân tương đối hiếm gặp gây mơ hồ về giới tính ở những nam giới có nhiễm sắc thể bình thường (Miller, 2002). Sự điều hòa ngược âm tính đối với sự sản xuất testosterone cũng xảy ra thông qua các estrogen và androgen (Darney et al., 1996; Ewing, 1983).
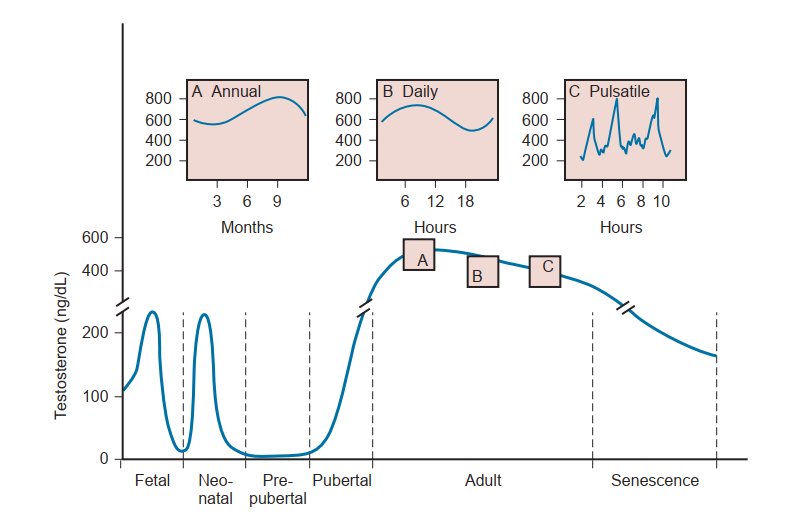
Các chu kỳ Testosterone. Nồng độ testosterone trong máu thay đổi mạnh mẽ trong suốt cuộc đời thai nhi, sơ sinh và trưởng thành của con người. Hình 64.9 cho thấy một đỉnh testosterone xảy ra ở thai nhi người vào tuần thứ 12 đến 18 của thai kỳ. Một đỉnh testosterone khác xảy ra vào khoảng 2 tháng tuổi. Một đỉnh testosterone thứ ba xảy ra trong thập kỷ thứ hai của cuộc đời. Sau đó, có một giai đoạn bình ổn (plateau), và sau đó là sự suy giảm chậm theo tuổi tác. Đan xen vào đó, có các nhịp điệu hàng năm và hàng ngày của sự sản xuất testosterone (xem Hình 64.9, hình nhỏ A và B) và các dao động không đều hàng ngày của testosterone (xem Hình 64.9, hình nhỏ C). Những thay đổi theo thời gian này trong sản xuất testosterone phản ánh một sự tương tác phức tạp và liên tục giữa tuyến yên và tinh hoàn. Các đỉnh testosterone tương ứng theo thời gian với bốn sự kiện phát triển: (1) sự phát triển của đường sinh dục thai nhi, (2) sự "in vết" ở trẻ sơ sinh của các mô đích phụ thuộc androgen, (3) sự nam tính hóa ở tuổi dậy thì, và (4) sự duy trì tăng trưởng và chức năng của các cơ quan phụ thuộc androgen ở người trưởng thành. Chủ đề này đã được Swerdloff và Heber (1981) tổng quan một cách toàn diện.
Các ống sinh tinh
Các ống sinh tinh bao gồm các tế bào mầm và các tế bào nâng đỡ, tạo thành một môi trường độc nhất cho sự sản xuất giao tử. Các tế bào nâng đỡ bao gồm tế bào Sertoli cùng các tế bào sợi và tế bào dạng cơ (myoid) của màng đáy. Các tế bào mầm bao gồm một quần thể tế bào gốc phân chia chậm, các tinh nguyên bào và tinh bào tăng sinh nhanh hơn, cùng các tinh tử đang biến thái.
Tế bào Sertoli. Các ống sinh tinh được lót bằng các tế bào Sertoli nằm trên màng đáy của ống và kéo dài các nhánh tế bào chất vào trong lòng ống (Hình 64.10). Các đặc điểm siêu cấu trúc của tế bào Sertoli đã được mô tả rõ ràng (Bardin et al., 1994). Chúng có các nhân hình dạng không đều, các hạch nhân nổi rõ, chỉ số phân bào thấp và thể hiện các phức hợp liên kết chặt (tight junctional complexes) độc đáo giữa các tế bào Sertoli liền kề. Những liên kết chặt này là những hàng rào giữa các tế bào mạnh nhất trong cơ thể. Chúng chia không gian ống sinh tinh thành các khoang đáy (màng đáy) và khoang cận lòng (lòng ống) (xem Hình 64.10). Sự sắp xếp giải phẫu này tạo cơ sở cho hàng rào máu - tinh hoàn và cho phép quá trình sinh tinh diễn ra ở một vị trí được đặc quyền về mặt miễn dịch. Các tế bào Sertoli đóng vai trò là các tế bào nuôi dưỡng (nurse cells) cho quá trình sinh tinh, nuôi dưỡng các tế bào mầm đang phát triển bên trong và giữa các nhánh bào tương của tế bào Sertoli. Các tinh nguyên bào chưa biệt hóa nằm gần màng đáy của ống, trong khi các tinh bào và tinh tử tiến hóa hơn nằm gần bề mặt lòng ống. Do đó, tế bào Sertoli là một biểu mô phân cực, trong đó phần đáy tiếp cận với môi trường huyết tương, và phần đỉnh của nó chứa một môi trường độc nhất đối với ống sinh tinh (Ewing et al., 1980).
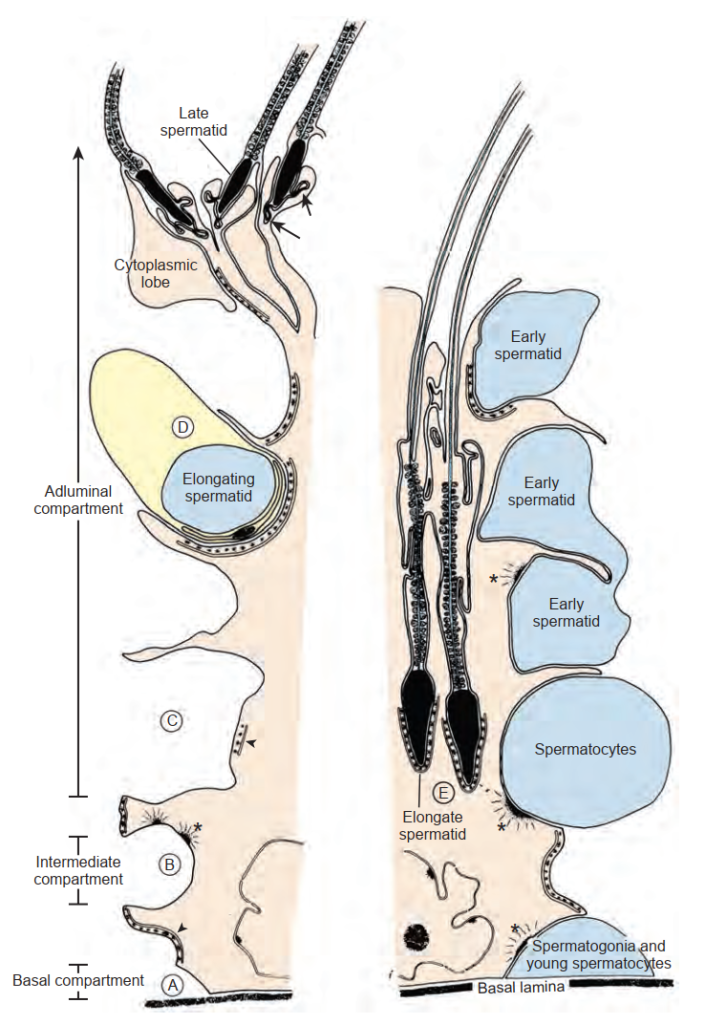
Các tế bào Sertoli nuôi dưỡng sự phát triển của tế bào mầm bằng cách (1) cung cấp một vi môi trường cận lòng chuyên biệt, (2) nâng đỡ các tế bào mầm thông qua các liên kết khe (gap junctions) giữa tế bào Sertoli và tế bào mầm, và (3) cho phép sự di cư của các tế bào mầm đang phát triển bên trong ống (xem Hình 64.10). Các liên kết chặt giữa các tế bào Sertoli liên tục được tái cấu trúc để cho phép sự "mở" và "đóng" cần thiết cho sự tương tác và di cư của tế bào mầm (Mruk và Cheng, 2004). Các phức hợp ligand - thụ thể, chẳng hạn như c-kit và kit ligand, có khả năng tham gia vào việc trung gian giao tiếp giữa tế bào mầm và tế bào Sertoli. Tế bào Sertoli cũng tham gia vào quá trình thực bào tế bào mầm, đồng thời sản xuất và bài tiết dịch cùng các phân tử hiệu ứng quan trọng. Protein liên kết androgen (ABP) là một trong những sản phẩm bài tiết của tế bào Sertoli được mô tả sớm nhất (Hansson và Djoseland, 1972). ABP là một chất vận chuyển nội bào của androgen bên trong tế bào Sertoli. Bằng cách liên kết với testosterone, ABP duy trì nồng độ androgen ở mức cao (cao gấp 50 lần so với huyết thanh) bên trong các ống sinh tinh. Testosterone cũng đóng một vai trò quan trọng trong sự điều hòa chức năng tế bào Sertoli, bao gồm cả sự sản xuất ABP (Griswold et al., 1988). Inhibin có nguồn gốc từ tế bào Sertoli và đóng một vai trò điều hòa quan trọng trong vòng điều hòa ngược âm tính của sự bài tiết FSH. Inhibin B là một dấu ấn nội tiết quan trọng của chức năng tế bào Sertoli trong việc đánh giá vô sinh nam (Pierik et al., 1998).
Với tư cách là những người bảo vệ thánh đường miễn dịch của tinh hoàn, các tế bào Sertoli duy trì một vi môi trường tế bào mầm hoàn toàn khác biệt so với huyết tương. Như vậy, tế bào Sertoli tiết ra nhiều sản phẩm khác, bao gồm các thành phần cơ chất ngoại bào (lamin, collagen týp IV, và collagen týp I) và các protein như ceruloplasmin, transferrin, glycoprotein 2, chất kích hoạt plasminogen, các chất giống somatomedin, protein T, kháng nguyên H-Y, clusterin, các protein vòng, các yếu tố tăng trưởng, và somatomedin (Mruk và Cheng, 2004). Dihydrotestosterone, testosterone, androstenediols, 17beta-estradiol, và các steroid C21 khác cũng được sản xuất bởi các tế bào Sertoli (Ewing et al., 1980; Mather et al., 1983), mặc dù vai trò của chúng trong chức năng tế bào Sertoli phần lớn vẫn chưa được biết đến.
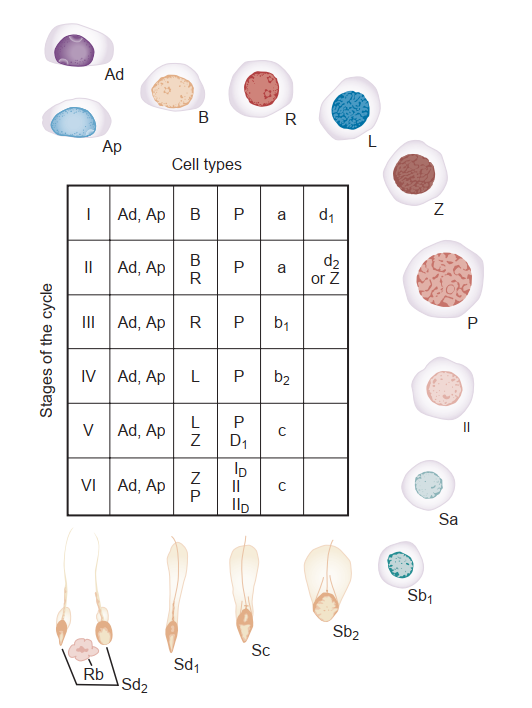
Tế bào mầm. Bên trong ống sinh tinh người, các tế bào mầm tạo ra khoảng 123 x 10^6 (khoảng dao động từ 21 đến 374 x 10^6) tinh trùng mỗi ngày (Amann và Howards, 1980). Điều này tương đương với việc sản xuất khoảng 1000 tinh trùng theo mỗi nhịp tim. Bên trong ống sinh tinh, các tế bào mầm được sắp xếp theo một trình tự có trật tự cao từ màng đáy đến lòng ống. Phân tích hình thái của các tế bào mầm khác nhau cho thấy có ít nhất 13 loại tế bào mầm có thể nhận biết được trong tinh hoàn người (Clermont, 1963; Heller và Clermont, 1964; Hình 64.11). Mỗi loại tế bào được cho là đại diện cho một bước khác nhau trong quá trình sinh tinh. Tiến triển từ ít biệt hóa nhất đến biệt hóa nhất, dựa trên hình thái xuất hiện, chúng được đặt tên là: tinh nguyên bào A tối (Ad); tinh nguyên bào A nhạt (Ap); tinh nguyên bào B (B); tinh bào một sơ cấp giai đoạn tiền leptotene (R), leptotene (L), zygotene (Z), và pachytene (P); tinh bào hai thứ cấp (II); và các tinh tử Sa, Sb, Sc, Sd1, và Sd2. Các liên kết chặt của tế bào Sertoli giữ các tinh nguyên bào và tinh bào giai đoạn sớm bên trong khoang đáy, và tất cả các tế bào mầm tiếp theo bên trong khoang cận lòng.
Cấu trúc quanh ống
Ống sinh tinh người được bao bọc bởi vài lớp mô quanh ống (Hermo et al., 1977; Hình 64.12). Lớp vỏ ngoài bao gồm các tế bào sợi. Ở lớp giữa là các tế bào dạng cơ xếp xen kẽ với các lá mô liên kết. Lớp trong cùng bao gồm một chất nền collagen. Các tế bào dạng cơ quanh ống được cho là có chức năng co bóp (Toyama, 1977). Các tế bào dạng cơ cũng tích cực tiết ra các thành phần chất nền ngoại bào là fibronectin và collagen týp I, đồng thời tạo ra lớp collagen trong cùng (Tung et al., 1984). Đáng chú ý, các tế bào dạng cơ được biết là liên kết với các tế bào Sertoli trong một sự tương tác trung mô - biểu mô chính xác. Skinner et al. (1988) đã phân lập được một yếu tố cận tiết có nguồn gốc từ tế bào cơ được sản xuất, đặt tên là P-Mod-S (peritubular modifies Sertoli - yếu tố quanh ống sửa đổi Sertoli), chất này ảnh hưởng sâu sắc đến các chức năng tổng hợp của tế bào Sertoli trong điều kiện in vitro.
Hàng rào Máu - Tinh hoàn
Các chất nhuộm và các chất khác, khi được tiêm vào dòng máu của động vật, nhanh chóng xuất hiện khắp tất cả các mô cơ thể nhưng không thể thâm nhập vào các vùng của não và tinh hoàn. Điều này dẫn đến khái niệm về sự tồn tại của một hàng rào máu - tinh hoàn. Tên gọi phù hợp hơn là "hàng rào máu - ống sinh tinh", nó có hai thành phần: một yếu tố giải phẫu hoặc cơ học và các yếu tố chức năng. Hàng rào cơ học được tạo ra, một phần, bởi các tế bào dạng cơ giống như cơ bao quanh các ống sinh tinh (Dym và Fawcett, 1970; Fawcett et al., 1970). Sự điều hòa sự di chuyển của các phân tử cũng xảy ra ở cấp độ của các tế bào nội mạc mao mạch. Tuy nhiên, thành phần quan trọng nhất của hàng rào này là các liên kết chặt khớp thần kinh (synaptic tight junctions) giữa các tế bào Sertoli nhằm ngăn chặn sự đi qua của các phân tử lớn và các tế bào lympho. Những yếu tố giải phẫu này là cần thiết nhưng chưa đủ để duy trì trạng thái "thánh đường" miễn dịch bên trong ống, bởi vì chúng không được quan sát thấy ở các vùng được bảo vệ khác của đường sinh sản (Brown et al., 1972; Tung et al., 1971).
Do đó, mặc dù hàng rào cơ học đóng góp vào sự cô lập của tinh hoàn, các thành phần "chức năng" khác cũng phải tồn tại để đàn áp phản ứng miễn dịch bình thường. Vài cơ chế có khả năng phối hợp với nhau để bảo vệ tinh trùng khỏi bị phá hủy. Đầu tiên, các loại và số lượng tế bào lympho bị giới hạn ở các vùng dễ bị tổn thương về mặt giải phẫu trong biểu mô mầm (Anderson và Hill, 1988; el-Demiry et al., 1985; Mahi-Brown et al., 1988). Cũng có bằng chứng cho thấy sự dung nạp miễn dịch đóng một vai trò trong hàng rào máu - tinh hoàn chức năng. Lý thuyết dẫn đầu đề xuất rằng bên trong các vùng yếu hơn về mặt giải phẫu (lưới tinh hoàn, ống xuất, mào tinh hoàn) của hàng rào, có một sự rò rỉ nhỏ, liên tục của các kháng nguyên tinh trùng (Tung, 1980). Sự rò rỉ này tạo ra các tế bào T-ức chế và sự dung nạp miễn dịch, tương tự như các phác đồ giải mẫn cảm đối với các chất gây dị ứng môi trường thông thường. Tuy nhiên, với những thách thức kháng nguyên lớn hơn, một phản ứng miễn dịch thực sự sẽ xảy ra (Turek, 1997). Đóng góp vào sự dung nạp này, giờ đây rõ ràng là các tế bào Sertoli biểu hiện các chất trung gian khác nhau hoạt động tại chỗ để tạo ra một môi trường ức chế miễn dịch bên trong tinh hoàn (Filippini et al., 2001). Chúng bao gồm phân tử "chất điều hòa miễn dịch bậc thầy" galectin 1 (Chui et al., 2011) cũng như các cytokine interferon-gamma, thụ thể Fc hòa tan, và TGF-beta (Ben-Rafael và Orvieto, 1992; Perussia et al., 1987; Turek, 1997), cùng các androgen (Diemer et al., 2003).
Tại sao hàng rào máu - tinh hoàn lại tồn tại? Bởi vì nó phát triển vào thời kỳ khởi phát sinh tinh (spermarche) (Kormano, 1967), nó có khả năng quan trọng đối với quá trình giảm phân, vì nó có thể cô lập về mặt miễn dịch các giao tử nam đang phát triển vốn không được hệ miễn dịch của người nam trưởng thành nhận diện là "vật tự thân" (self). Theo nghĩa này, giá trị của hàng rào máu - tinh hoàn được hiện thực hóa đầy đủ sau tuổi dậy thì, bởi vì các "kháng nguyên" ngoại lai trên các tế bào mầm sau giảm phân chỉ tồn tại sau thời kỳ khởi phát sinh tinh. Một tổn thương tinh hoàn như sinh thiết, xoắn, hoặc chấn thương sẽ không kích ứng các kháng thể kháng tinh trùng nếu nó xảy ra trước tuổi dậy thì. Tuy nhiên, sau tuổi dậy thì, vô sinh do miễn dịch là một nguy cơ đã được biết đến (Turek, 1997). Trên lâm sàng, hàng rào máu - tinh hoàn cũng có thể hạn chế sự tiếp cận của hóa trị liệu đến các tế bào ung thư đang lẩn trốn phía sau nó và dẫn đến sự tái phát ung thư cô lập bên trong tinh hoàn.
Quá trình sinh tinh
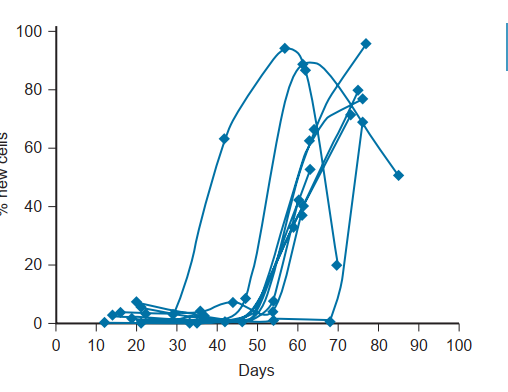
Quá trình sinh tinh là một tiến trình đặc biệt phức tạp và chuyên biệt của sự giảm giảm phân DNA và sự biến thái của tế bào mầm. Các nghiên cứu cũ hơn đã ước tính rằng toàn bộ quá trình ở người đòi hỏi khoảng 64 ngày (Clermont, 1972). Tuy nhiên, một nghiên cứu động học in vivo ở những nam giới khỏe mạnh đã tiết lộ rằng tổng thời gian để sản xuất ra một tinh trùng được phóng tinh dao động từ 42 đến 76 ngày, gợi ý rằng thời gian sinh tinh có thể thay đổi rộng rãi giữa các cá thể (Misell et al., 2006; Hình 64.13). Quá trình sinh tinh bao gồm (1) một giai đoạn tăng sinh khi các tinh nguyên bào phân chia để thay thế số lượng của chúng (tự làm mới) hoặc biệt hóa thành các tế bào con để trở thành các giao tử trưởng thành; (2) một giai đoạn giảm phân khi các tế bào mầm trải qua một sự phân chia giảm nhiễm, dẫn đến các tinh tử đơn bội (chứa một nửa bộ DNA bình thường); và (3) một giai đoạn tạo tinh trùng (spermiogenesis) trong đó các tinh tử trải qua một sự biến thái sâu sắc để trở thành các tinh trùng trưởng thành. (Để có các bài tổng quan xuất sắc, xem Steinberger [1976] và de Kretser và Kerr [1988].)
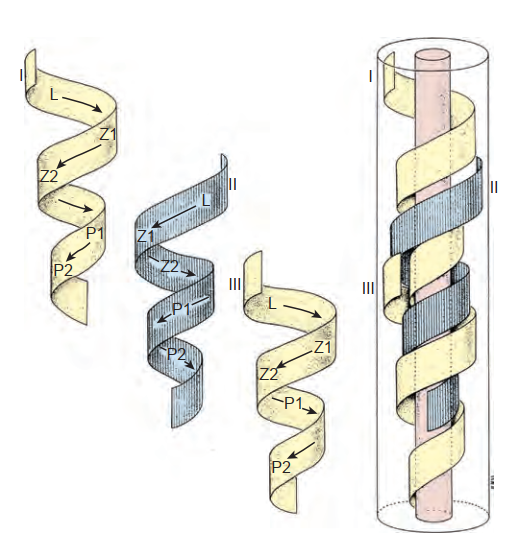
Một chu kỳ sinh tinh liên quan đến sự phân chia của các tế bào gốc tinh nguyên bào nguyên thủy thành các tế bào mầm tiếp theo. Vài chu kỳ sinh tinh cùng tồn tại bên trong biểu mô mầm, và chúng được mô tả về mặt hình thái như là các giai đoạn (stages). Nếu quá trình sinh tinh được quan sát từ một điểm cố định duy nhất bên trong một ống sinh tinh, sáu tổ hợp tế bào hoặc giai đoạn có thể nhận biết được quan sát thấy một cách dự đoán trước ở người (Heller và Clermont, 1964) (xem Hình 64.11). Ngoài ra, cũng có một sự tổ chức cụ thể của các chu kỳ sinh tinh bên trong không gian ống, được gọi là các sóng sinh tinh (spermatogenic waves). Bằng chứng tốt nhất cho thấy quá trình sinh tinh ở người tồn tại dưới một sự sắp xếp tế bào theo hình xoắn ốc hoặc đinh ốc nhằm đảm bảo sự sản xuất tinh trùng là một quá trình liên tục chứ không phải là một quá trình theo mạch xung (Schulze, 1989; Hình 64.14).
Sự di cư, tự làm mới và tăng sinh của Tế bào gốc Tinh hoàn
Sự di cư của Tế bào gốc Tinh hoàn. Trong quá trình phát triển sớm trước khi sinh, các tế bào mầm nguyên thủy di cư đến gờ sinh dục và liên kết với các tế bào Sertoli để hình thành các dây tinh hoàn nguyên thủy (Witschi, 1948).
Những tế bào gốc dòng mầm nguyên thủy này được gọi là các tế bào sinh dục đầu dòng (gonocytes) sau khi tuyến sinh dục biệt hóa thành tinh hoàn bằng cách hình thành các dây sinh tinh. Chúng được gọi là tinh nguyên bào sau khi di cư đến vùng ngoại vi của ống (Gondos và Hobel, 1971). Những tế bào mầm di cư giai đoạn sớm này có các đặc tính tương tự như các tế bào gốc phôi và có khả năng là nguồn gốc của các khối u tế bào mầm ở người trưởng thành (Ezeh et al., 2005). Sự thất bại của các tế bào mầm trong việc di cư vào tinh hoàn nguyên thủy cũng được cho là nguyên nhân gây ra các khối u tế bào mầm ngoài tuyến sinh dục và chứng vô sinh ở người trưởng thành do hội chứng chỉ có tế bào Sertoli (Sertoli cell–only) trên mô bệnh học tinh hoàn gây ra vô tinh (Nikolic et al., 2016).
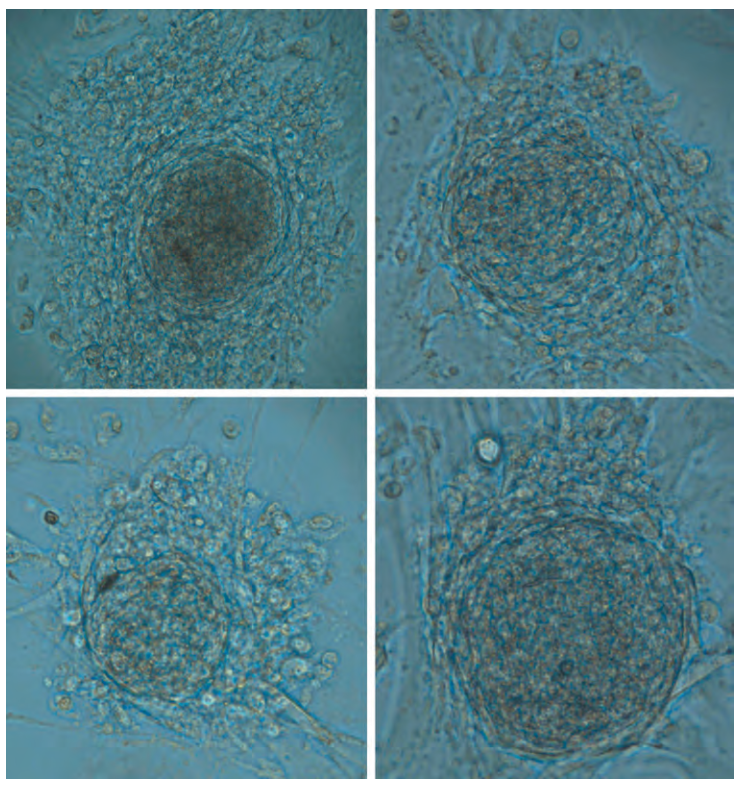
Sự tự làm mới của Tế bào gốc Tinh hoàn (Testis Stem Cell Renewal). Các tinh nguyên bào nằm trong hốc tế bào gốc tinh hoàn (testis stem cell niche) được bổ sung liên tục thông qua một tiến trình gọi là sự tự làm mới tế bào gốc. Hệ thống thụ thể kit ligand/c-kit receptor (yếu tố phát triển tế bào gốc) và yếu tố vùng hốc là yếu tố dinh dưỡng thần kinh có nguồn gốc từ dòng tế bào thần kinh đệm (GDNF) dường như có tham gia vào tiến trình này (Oatley và Brinster, 2008). Các nghiên cứu gần đây cũng chỉ ra rằng các tế bào gốc tinh nguyên bào ở người có thể được tái lập trình trong điều kiện in vitro để trở thành các tế bào gốc giống phôi (embryonic-like stem cells) (Conrad et al., 2008; Kossack et al., 2009; Hình 64.15). Được thu thập từ các mẫu sinh thiết tinh hoàn của người trưởng thành, các tế bào giống phôi này biểu hiện các dấu ấn vạn năng đặc trưng (OCT-4, SOX-2, STELLAR, GDF-3), có thể biệt hóa thành cả ba lá phôi, duy trì một bộ nhiễm sắc thể (karyotype) bình thường, có khả năng hình thành u quái (teratomas), và biểu hiện các dấu ấn biểu sinh cùng enzyme telomerase ở mức độ phù hợp (Kossack et al., 2009). Phát hiện này gợi ý rằng trong tương lai, tinh hoàn có thể trở thành một nguồn cung cấp các tế bào gốc đặc hiệu cho từng bệnh nhân phục vụ cho các liệu pháp điều trị dựa trên tế bào.
Sự tăng sinh của Tế bào gốc Tinh hoàn (Testis Stem Cell Proliferation). Ở người, các tinh nguyên bào A nhạt (Ap) nằm trong hốc tế bào gốc vùng đáy của ống sinh tinh sẽ phân chia theo chu kỳ mỗi 16 ngày (Clermont, 1972) để hình thành nên các tinh nguyên bào B. Các tinh nguyên bào B được định hướng để trở thành các tinh bào, nhưng phần tế bào chất giữa các tế bào tinh nguyên bào con vẫn giữ nguyên sự kết nối sau quá trình nguyên phân, tạo thành các cầu tế bào chất (cytoplasmic bridges) giữa các tế bào liền kề. Những cầu tế bào chất này được quan sát thấy giữa các tế bào mầm thuộc mọi phân lớp trong suốt quá trình sinh tinh (Ewing et al., 1980). Các cầu nối này có thể đóng vai trò quan trọng đối với sự tăng sinh và biệt hóa tế bào mang tính đồng bộ, cũng như đối với việc điều hòa biểu hiện gen.
Giảm phân (Meiosis)
Các tế bào sinh dưỡng (somatic cells) nhân đôi bằng quá trình nguyên phân (mitosis), tạo ra các tế bào con có đặc điểm di truyền giống hệt nhau. Các tế bào mầm (germ cells) nhân đôi bằng quá trình giảm phân (meiosis), trong đó vật chất di truyền bị giảm đi một nửa để phục vụ cho sự sinh sản. Quá trình giảm phân tạo ra sự đa dạng di truyền, cung cấp một nguồn vật liệu phong phú hơn để chọn lọc tự nhiên có thể tác động lên. Sự nhân đôi tế bào bằng nguyên phân là một chuỗi các sự kiện diễn ra chính xác, được phối hợp nhịp nhàng bao gồm sự sao chép vật chất di truyền (nhiễm sắc thể), sự tiêu biến của màng nhân, cùng sự phân chia đồng đều của các nhiễm sắc thể và tế bào chất vào các tế bào con. Sự khác biệt căn bản giữa sự nhân đôi nguyên phân và giảm phân là: ở quá trình nguyên phân, một bước sao chép DNA duy nhất chỉ được theo sau bởi một lần phân chia tế bào; còn ở quá trình giảm phân, bước sao chép đó được theo sau bởi hai lần phân chia tế bào (tạo ra bốn tế bào con). Hệ quả là, các tế bào con chỉ chứa một nửa hàm lượng nhiễm sắc thể của tế bào mẹ. Do đó, một tế bào mẹ lưỡng bội (2n) sẽ trở thành một giao tử đơn bội (n). Các khác biệt lớn khác giữa nguyên phân và giảm phân được tóm tắt trong Bảng 64.2. Nghiên cứu đã chỉ ra rằng các phân tử RNA kích thước nhỏ (small RNAs), bao gồm các RNA can thiệp nhỏ (siRNAs), microRNAs (miRNAs), và các RNA tương tác với piwi (piRNAs), là những chất điều hòa quan trọng đối với sự biểu hiện gen của tế bào mầm ở cấp độ sau phiên mã hoặc cấp độ dịch mã (He et al., 2009; Tolia và Joshua-Tor, 2007).
BẢNG 64.2 Các khác biệt căn bản: Nguyên phân và Giảm phân
| NGUYÊN PHÂN (MITOSIS) | GIẢM PHÂN (MEIOSIS) |
| Xảy ra ở các tế bào sinh dưỡng | Xảy ra ở các tế bào sinh dục |
| Một lần phân chia tế bào, tạo ra hai tế bào con | Hai lần phân chia tế bào, tạo ra bốn tế bào con |
| Số lượng nhiễm sắc thể được duy trì | Số lượng nhiễm sắc thể giảm đi một nửa |
| Không có sự bắt cặp của các nhiễm sắc thể tương đồng | Có sự tiếp hợp (synapse) của các nhiễm sắc thể tương đồng ở kỳ đầu I |
| Không có sự trao đổi chéo (crossovers) | Có nhiều hơn một sự trao đổi chéo trên mỗi cặp nhiễm sắc thể tương đồng |
| Tâm động phân chia ở kỳ sau | Tâm động phân chia ở kỳ sau II |
| Kiểu gen của tế bào con giống hệt nhau | Có sự biến dị di truyền ở các tế bào con |
Quá trình sinh tinh bắt đầu với việc các tinh nguyên bào týp B phân chia nguyên phân để hình thành các tinh bào một sơ cấp (primary spermatocytes) bên trong khoang cận lòng. Các tinh bào một sơ cấp là những tế bào mầm đầu tiên trải qua quá trình giảm phân (Kerr và de Kretser, 1981). Trong tiến trình này, một lần phân chia giảm phân được theo sau bởi một lần phân chia giảm nhiễm kiểu nguyên phân điển hình, kết quả tạo ra các tế bào con có một bộ nhiễm sắc thể đơn bội. Ngoài ra, như một hệ quả của sự tái tổ hợp nhiễm sắc thể, mỗi tế bào con sẽ chứa đựng những thông tin di truyền khác nhau. Tế bào thu được sau tiến trình này là tinh tử Sa (Sa spermatid) (xem Hình 64.11).
Sự tái tổ hợp nhiễm sắc thể (chromosomal recombination), đặc trưng định nghĩa của quá trình giảm phân ở động vật có vú, đảm bảo rằng các giao tử đơn bội có sự khác biệt về mặt di truyền so với các tế bào tiền thân trưởng thành của chúng, và đây chính là động cơ thực sự của sự đa dạng di truyền và tiến hóa. Trong kỳ đầu của giảm phân, sự hình thành một phức hợp tiếp hợp (synaptonemal complex) cùng với sự bắt cặp của các nhiễm sắc thể tương đồng (nguồn gốc từ mẹ và từ cha) sẽ diễn ra, đi kèm với sự tương tác vật lý và trao đổi DNA thông qua các vị trí trao đổi chéo tương hỗ (chiasmata) giữa các nhiễm sắc thể tương đồng. Nghiên cứu gần đây đã chỉ ra rằng các khiếm khuyết về độ chính xác của sự tái tổ hợp bên trong các tế bào mầm nam giới ở người có thể gây ra tình trạng vô tinh và vô sinh nam (Walsh et al., 2009). Trong một nghiên cứu, có 10% nam giới bị vô tinh không bế tắc (nonobstructive azoospermia) mang những khiếm khuyết đáng kể trong quá trình tái tổ hợp so với những nam giới có quá trình sinh tinh bình thường (Gonsalves et al., 2004). Ngoài ra, trong số những người đàn ông có kiểu hình ngừng trưởng thành (maturation arrest) trên kết quả sinh thiết tinh hoàn, sự tái tổ hợp lỗi được quan sát thấy ở khoảng một nửa số trường hợp, cung cấp bằng chứng cho thấy sự tái tổ hợp lỗi có mối liên hệ với tình trạng sản xuất tinh trùng kém (Gonsalves et al., 2004). Những biến đổi trong tái tổ hợp cũng có những ảnh hưởng đến tình trạng dị bội của tinh trùng, bởi vì các thay đổi ở vị trí trao đổi chéo là những yếu tố nguy cơ dẫn đến sự không phân ly nhiễm sắc thể (chromosomal nondisjunction). Thật vậy, các bằng chứng gợi ý rằng mối tương quan giữa sự tái tổ hợp lỗi và tình trạng dị bội của tinh trùng ở nam giới bị vô tinh là đủ mạnh để giải thích cho tỷ lệ bất thường nhiễm sắc thể cao hơn ở thế hệ con được thụ tinh bằng phương pháp thụ tinh trong ống nghiệm (IVF) và tiêm tinh trùng vào bào tương noãn (ICSI) (Sun et al., 2008).
Quá trình tạo tinh trùng / Biến thái tinh tử (Spermiogenesis)
Trong quá trình tạo tinh trùng, các tinh tử Sa hình tròn sẽ trưởng thành để trở thành các tinh trùng (spermatose) (xem Hình 64.11). Trong chuỗi trưởng thành này, sự phân chia tế bào không diễn ra, nhưng có những thay đổi sâu rộng đối với nhân và tế bào chất của tinh tử. Chúng bao gồm việc mất tế bào chất, sự di cư của các bào quan tế bào chất, sự hình thành cực đầu (acrosome) từ bộ máy Golgi, sự hình thành roi (flagellum) từ trung tử, sự cô đặc nhân xuống còn khoảng 10% kích thước trước đây, và sự sắp xếp lại của các ty thể xung quanh đoạn giữa của tinh trùng (Kerr và de Kretser, 1981). Nhân của tinh tử tròn chuyển từ hình cầu sang hình bất đối xứng khi chất nhiễm sắc cô đặc lại. Nhiều yếu tố tế bào đóng góp vào tiến trình tái định hình này, bao gồm cấu trúc nhiễm sắc thể, các protein nhiễm sắc thể liên kết, lớp vỏ khung tế bào quanh nhân (perinuclear cytoskeletal theca layer), bao vi ống của nhân (manchette of nuclear microtubules), actin dưới cực đầu, và các tương tác của tế bào Sertoli. Khi quá trình kéo dài của tinh tử hoàn tất, tế bào chất của tế bào Sertoli sẽ co rút lại xung quanh tinh trùng đang phát triển, tước bỏ toàn bộ phần tế bào chất không cần thiết của nó và tống xuất nó vào trong lòng ống sinh tinh. Tinh trùng trưởng thành có lượng tế bào chất ít một cách đáng kinh ngạc và được sản xuất với số lượng khổng lồ — lên đến 300 tinh trùng trên mỗi gam tinh hoàn trong một giây.
Sự tương tác giữa Tế bào Sertoli và Tế bào mầm
Một mạng lưới tương tác tế bào - tế bào phức tạp tồn tại bên trong tinh hoàn giữa các tế bào Leydig và tế bào Sertoli, giữa các tế bào Leydig và tế bào quanh ống, giữa tế bào Sertoli và tế bào quanh ống, và giữa tế bào Sertoli với các tế bào mầm. Một số tổ hợp tế bào Sertoli – tế bào mầm ở tinh hoàn động vật có vú được minh họa trong Hình 64.10 (Romrell và Ross, 1979; Russell và Clermont, 1976; Skinner, 1995). Ngoài ra, còn có các yếu tố có thể phá vỡ hàng rào máu - tinh hoàn một cách thuận nghịch, bao gồm TGF-beta3 và yếu tố hoại tử u alpha(TNF-alpha). Các chất này hoạt động bằng cách làm giảm nồng độ của occludin và zonula occludens-1 (ZO-1) trong hàng rào thông qua con đường truyền tín hiệu mitogen-activated protein (MAP) kinase p38 (Xia et al., 2009). Điều này chỉ đại diện cho một phần của tiến trình cực kỳ phức tạp và có tính tương tác cao đặc trưng cho quá trình sinh tinh.
Di truyền học
Các nguyên nhân di truyền gây bất thường quá trình sinh tinh đã được xác định dưới dạng các đột biến điểm ở các gen đơn lẻ di truyền theo kiểu Mendelian (ví dụ: xơ nang), và dưới dạng các rối loạn nhiễm sắc thể trong đó các đoạn của nhiễm sắc thể (hoặc toàn bộ nhiễm sắc thể) có những bất thường về mặt cấu trúc hoặc số lượng. Độc giả được giới thiệu đến tài liệu của Turek và Reijo Pera (2002) để có một bài tổng quan toàn diện về các rối loạn này. Giả thuyết cho rằng các đoạn mất đoạn ở nhánh dài của nhiễm sắc thể Y gây ra tình trạng vô tinh đã được đưa ra cách đây hơn ba thập kỷ (Tiepolo et al., 1976). Dựa trên phân tích di truyền tế bào, vùng mang tính lý thuyết này được đặt tên là yếu tố gây vô tinh (AZF). Hiện tại, các kiểu vị trí của các mất đoạn (được gọi là mất đoạn nhỏ / microdeletions) trong vùng AZF được sử dụng để chia vùng này thành các vùng phụ là AZFa, AZFb, và AZFc (Vogt et al., 1996). Các mất đoạn vùng của nhiễm sắc thể Y, được gọi là mất đoạn nhỏ Yq, xảy ra ở 6% đến 8% nam giới bị thiểu tinh nặng và ở 15% nam giới bị vô tinh (Reijo et al., 1996). Nhìn chung, những mất đoạn như vậy là nguyên nhân phân tử được xác định phổ biến nhất của vô sinh nam (Kostiner et al., 1998).
Đang có các tài liệu y văn mới đề cập đến giá trị tiên lượng của các mất đoạn AZF. Trái ngược với những bệnh nhân mất đoạn AZFc một phần và hoàn toàn, những người thường vẫn tìm thấy tinh trùng khi phân tích tinh dịch hoặc sinh thiết tinh hoàn, việc tìm thấy tinh trùng trong tinh dịch hoặc trong tinh hoàn ở nam giới có mất đoạn hoàn toàn AZFa hoặc AZFb là điều cực kỳ khó xảy ra (Hopps et al., 2003; Park et al., 2013). Mất đoạn AZFa hoàn toàn có liên quan đến tình trạng bất sản tế bào mầm hoặc hình thái chỉ có tế bào Sertoli (Sertoli cell–only). Nói chung, mất đoạn AZFb hoàn toàn có liên quan đến sự ngừng trưởng thành ở các giai đoạn tinh bào một sơ cấp (sớm) hoặc tinh tử (muộn), nhưng tinh trùng trong tinh dịch cũng đã được báo cáo trong một vài trường hợp cá biệt (Park et al., 2013). Mất đoạn AZFc có liên quan đến tình trạng giảm sinh tinh (hypospermatogenesis) hoặc kiểu hình chỉ có tế bào Sertoli đan xen với các ổ sinh tinh. Tinh trùng đã được phát hiện trong tinh dịch của những nam giới bị nghi ngờ và đã được xác nhận là có mất đoạn AZFa và AZFb một phần (Foresta et al., 2001). Tương tự, tinh trùng trong tinh dịch ở nam giới có mất đoạn AZFa + b, và AZFb + c (có thể là các mất đoạn một phần) cũng đã được báo cáo (Park et al., 2013), nhưng việc tìm thấy mất đoạn AZFa - c có liên quan đến tình trạng vô tinh và không có tinh trùng khi sinh thiết tinh hoàn.
Gần đây hơn, rõ ràng là nhiễm sắc thể X cũng quan trọng đối với quá trình sinh tinh. Vào năm 2001, Wang et al. đã báo cáo về một cuộc tìm kiếm hệ thống các gen chỉ biểu hiện ở tinh nguyên bào chuột và phát hiện thấy 10 gen nằm trên nhiễm sắc thể X (Wang et al., 2001). Các nghiên cứu sâu hơn về các gen liên kết nhiễm sắc thể X ở những bệnh nhân vô sinh nam đã xác định gen SOX3 (sex determining region Y box 3) và gen FATE là hai gen ứng viên tiềm năng cho khả năng sinh sản (Olesen et al., 2003; Raverot et al., 2004), xác nhận khái niệm rằng các gen trên nhiễm sắc thể X có thể đóng góp đáng kể vào các trường hợp vô sinh nam hiện chưa giải thích được (Mueller et al., 2013).
Di truyền học và Tuổi của người cha
Các bất thường nhiễm sắc thể tinh trùng liên quan đến tuổi. Tình trạng nhiễm sắc thể của tinh trùng lần đầu tiên được nghiên cứu do lo ngại rằng tuổi cao của người cha có liên quan đến sự gia tăng các trường hợp tam nhiễm (trisomy), đặc biệt là tam nhiễm 21 hoặc hội chứng Down ở thế hệ con. Với công nghệ lai tại chỗ phát huỳnh quang (FISH), những ảnh hưởng tinh tế của tuổi cha lên tình trạng dị bội (aneuploidy) của tinh trùng hiện đã rõ ràng. Ảnh hưởng của tuổi cha dường như làm tăng tỷ lệ tinh trùng bị dị bội nhiễm sắc thể giới tính (Wyrobek et al., 1996). Tuy nhiên, có rất ít bằng chứng hỗ trợ cho việc gia tăng các ca sinh dị bội liên quan đến tuổi cha, ngoại trừ có khả năng là tam nhiễm 21 và lưỡng nhiễm 1 (rất hiếm). Nghiên cứu các bất thường về cấu trúc nhiễm sắc thể tinh trùng, Martin và Rademaker (1987) đã mô tả một mối quan hệ tuyến tính có ý nghĩa giữa tuổi cha và tần suất các bất thường cấu trúc ở tinh trùng (r = 0,63). Một lời giải thích cho mối liên hệ này có thể là sự phân chia tế bào liên tục trong quá trình sinh tinh khiến các tế bào mầm đối mặt với nguy cơ tổn thương nhiễm sắc thể, đặc biệt là khi tuổi cha đã cao. Tuy nhiên, ngoại trừ các chuyển đoạn tương hỗ (reciprocal translocations), có rất ít bằng chứng chỉ ra rằng mối liên hệ này dẫn đến việc tăng tần suất con cái sinh ra có các bất thường cấu trúc nhiễm sắc thể tân phát (de novo).
Các đột biến gen tinh trùng liên quan đến tuổi. Các khiếm khuyết đơn gen ở tinh trùng là kết quả của các sai sót trong quá trình sao chép DNA. Mặc dù rất khó để đánh giá sự hiện diện hay vắng mặt của các khiếm khuyết đó trong tinh trùng, nhưng ảnh hưởng của tuổi cha cao đối với các bệnh lý ở thế hệ con có liên quan đến đột biến đơn gen là rất rõ ràng. Các rối loạn này được liệt kê trong Khung 64.1 và bao gồm các bệnh lý di truyền trội trên nhiễm sắc thể thường có mối liên hệ đã được biết đến với tuổi cha cao. Chúng được gọi là các kiểu hình lính gác (sentinel phenotypes) vì chúng là các rối loạn có tần suất đáng kể và độ thích nghi thấp (low fitness), đồng thời bắt nguồn từ các đột biến có độ đâm xuyên cao (highly penetrant). Một cơ chế cho sự phát triển của các đột biến đơn gen mới theo tuổi tác có liên quan đến tiến trình phân chia đặc trưng và liên tục của tế bào tinh nguyên bào. Đến tuổi dậy thì, đã có 30 lần phân chia tế bào tinh nguyên bào xảy ra; tuy nhiên, sau tuổi dậy thì, có 23 lần phân chia xảy ra mỗi năm ở các tế bào này. Sự thật đơn giản là các tinh nguyên bào của những người đàn ông lớn tuổi đã trải qua vô số lần phân chia tế bào có thể làm tăng cơ hội xảy ra các sai sót trong quá trình phiên mã DNA, nguồn gốc của các khiếm khuyết đơn gen. Các ước tính nguy cơ chính thức tồn tại đối với sự đóng góp của tuổi cha cao vào các đột biến trội trên nhiễm sắc thể thường. Ở nam giới dưới 29 tuổi, nguy cơ xảy ra đột biến ở thế hệ con là 0,22 trên 1000 ca sinh. Nguy cơ này tăng gấp đôi (0,45 trên 1000) ở độ tuổi cha từ 40 đến 44, và sau đó leo thang lên 3,7 trên 1000 ca sinh ở các độ tuổi trên 45 (Friedman, 1981).
Các thay đổi biểu sinh tinh trùng liên quan đến tuổi. Ngày càng có nhiều bằng chứng cho thấy các dấu ấn methyl hóa DNA trong tinh trùng giúp kiểm soát sự biểu hiện gen, được gọi là biểu sinh (epigenetics), bị thay đổi theo tuổi của người cha (Jenkins et al., 2014). Tinh trùng dường như tích tụ hàng trăm khiếm khuyết methyl hóa DNA theo tuổi tác, các khiếm khuyết này khu trú tại các vị trí bộ gen cụ thể, nhiều vị trí trong số đó kiểm soát các gen liên quan đến sự phát triển thần kinh (ví dụ: tâm thần phân liệt, bệnh lưỡng cực, tự kỷ, rối loạn tâm trạng). Trong một nghiên cứu về tinh trùng được hiến tặng hai lần bởi cùng một nhóm nam giới, một lần khi còn trẻ và một lần nữa khi đã lớn tuổi, tốc độ thay đổi biểu sinh trong tinh trùng cao gấp đôi so với tốc độ được ước tính cho các mô cơ thể khác (Jenkins et al., 2014). Thậm chí còn thú vị hơn, các thay đổi biểu sinh xảy ra ở tinh trùng khi nam giới già đi có xu hướng tập trung ở các gen liên quan đến bệnh tâm thần phân liệt và rối loạn lưỡng cực, những bệnh được biết là xảy ra nhiều hơn ở thế hệ con khi tuổi cha tăng lên. Do đó, ít nhất một nhóm nhỏ các rối loạn liên quan đến tuổi cha ở thế hệ con có thể là hệ quả của các biến đổi biểu sinh được truyền qua tinh trùng (Yatsenko và Turek, 2018).
KHUNG 64.1 Các rối loạn di truyền ở thế hệ con có liên quan đến tuổi cha cao
- Loạn sản sụn (Achondroplasias)
- Tật không mống mắt (Aniridia)
- Hội chứng Apert
- U nguyên bào võng mạc hai bên (Bilateral retinoblastoma)
- Hội chứng Crouzon
- Loạn sản sợi hóa xương tiến triển (Fibrodysplasia ossificans)
- Bệnh máu khó đông A (Hemophilia A)
- Hội chứng Lesch-Nyhan
- Hội chứng Marfan
- Bệnh u xơ thần kinh (Neurofibromatosis)
- Hội chứng mắt - răng - ngón (Oculodentodigital syndrome)
- Bệnh thận đa nang (Polycystic kidney disease)
- Đa polyp đại trực tràng (Polyposis coli)
- Bệnh già trước tuổi (Progeria)
- Hội chứng Treacher Collins
- Bệnh xơ cứng củ (Tuberous sclerosis)
- Hội chứng Waardenburg
- Tâm thần phân liệt (Schizophrenia)
- Rối loạn lưỡng cực (Bipolar disorder)
- Tự kỷ (Autism)
- Các rối loạn thiếu chú ý (Attention deficit disorders)
- Bệnh Alzheimer (được giả thuyết)
CÁC ĐIỂM CHÍNH: TINH HOÀN
- Tinh hoàn chứa 250 m ống sinh tinh và 700 triệu tế bào Leydig ở người trẻ trưởng thành.
- Quá trình sinh tinh diễn ra theo các giai đoạn, chu kỳ và sóng để đảm bảo sự sản xuất tinh trùng liên tục.
- Các gen trên nhiễm sắc thể X, cũng như nhiễm sắc thể Y, chi phối quá trình sinh tinh và góp phần gây vô sinh nam.
- Theo tuổi của người cha, có sự gia tăng các bất thường cấu trúc nhiễm sắc thể tinh trùng, các đột biến trội trên nhiễm sắc thể thường, và các biến đổi biểu sinh dẫn đến bệnh tật ở thế hệ con.
MÀO TINH HOÀN
Cấu trúc đại thể
Mào tinh hoàn là một cơ quan hình dấu phẩy nằm dọc theo mặt sau bên của tinh hoàn. Việc đi qua mào tinh hoàn tạo ra nhiều thay đổi cho tinh trùng mới được hình thành, bao gồm việc đạt được khả năng di động chức năng, và các thay đổi về điện tích bề mặt, các protein màng, độ đáp ứng miễn dịch, các phospholipid, hàm lượng axit béo và hoạt tính adenylate cyclase. Những thay đổi này cải thiện tính toàn vẹn cấu trúc màng tế bào, tăng cường khả năng di động cùng tính hướng hóa của tinh trùng, và làm tăng khả năng thụ tinh. Các tinh trùng bên trong tinh hoàn có khả năng di động rất kém hoặc không di động. Chúng trở nên di động tiến tới và có chức năng chỉ sau khi đi qua mào tinh hoàn.
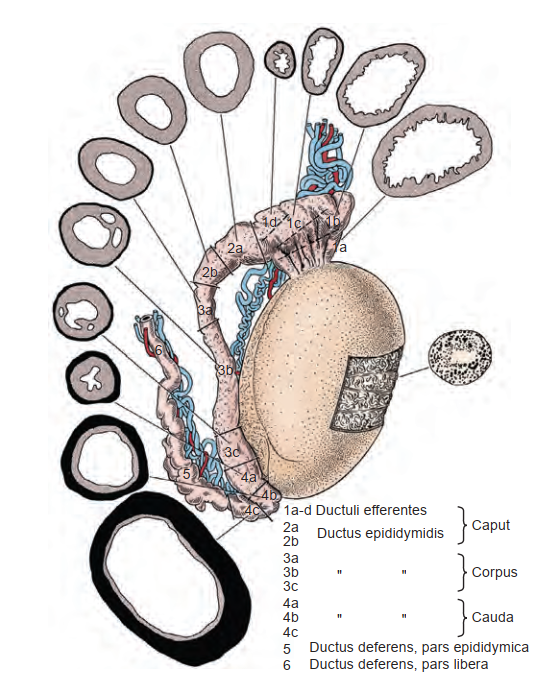
Mào tinh hoàn là một ống hoặc ống dẫn có chiều dài từ 3 đến 4 m, cuộn xoắn chặt chẽ và được bao bọc trong bao mô liên kết của màng tinh hoàn (tunica vaginalis) (Lanz và Neuhauser, 1964; Turner et al., 1978). Các nhánh từ bao xơ đi vào các khoảng gian ống và hình thành các vách ngăn chia ống thành các vùng có đặc điểm mô học riêng biệt (Kormano và Reijonen, 1976). Về mặt giải phẫu, chúng được chia theo kiểu cổ điển thành đầu mào tinh (caput), thân mào tinh (corpus), và đuôi mào tinh (cauda) (Hình 64.16). Đầu mào tinh hoàn bao gồm từ 8 đến 12 ống xuất đi ra từ tinh hoàn. Lòng của các ống xuất có kích thước lớn và hình dạng hơi không đều ở gần tinh hoàn, trở nên hẹp và có hình bầu dục ở gần vị trí nối với ống mào tinh. Phía xa của vị trí nối này, đường kính ống tăng lên một chút và sau đó duy trì không đổi ở phần thân mào tinh. Ở vùng đuôi mào tinh hoàn có kích thước lớn, đường kính ống tăng lên đáng kể và có hình dạng không đều. Tiến về phía xa, ống dần dần chuyển sang hình thái xuất hiện đặc trưng của ống dẫn tinh.
Nguồn cung cấp mạch máu và bạch huyết
Ở người, phần đầu và phần thân mào tinh hoàn nhận máu động mạch từ một nhánh của động mạch tinh hoàn (xem Hình 64.7). Nhánh này sau đó chia thành các nhánh mào tinh hoàn trên và dưới (MacMillan, 1954). Mào tinh hoàn cũng nhận máu từ các nhánh của động mạch ống dẫn tinh (deferential arteries), và các mạch máu bên kết nối động mạch ống dẫn tinh với nguồn cung cấp máu của tinh hoàn. Đuôi mào tinh hoàn được cung cấp bởi các nhánh của động mạch ống dẫn tinh. Động mạch ống dẫn tinh và động mạch cơ bìu phục vụ như các nguồn mạch máu bên cho mào tinh hoàn khi động mạch tinh hoàn chính bị tắc nghẽn hoặc bị thắt. Các nhánh động mạch bên trong mào tinh hoàn đi vào dọc theo các vách ngăn được hình thành từ bao mô liên kết. Các mạch máu này cuộn xoắn rất nhiều trước khi biến đổi thành các mạch máu thẳng của lưới vi tuần hoàn (Kormano và Reijonen, 1976). Mật độ vi mạch thay đổi đáng kể dọc theo chiều dài của mào tinh hoàn, với phần đầu gần chứa mạng lưới mao mạch dưới biểu mô dày đặc nhất và các đoạn xa hơn chứa hệ thống mạch máu ít dày đặc hơn. Từ các nghiên cứu trên động vật, mạng lưới mao mạch mào tinh hoàn chịu sự kiểm soát của hormone. Ví dụ, ở thỏ, việc thiến hormone hai bên dẫn đến sự thoái hóa tiến triển và cuối cùng là biến mất của mạng lưới mao mạch mào tinh hoàn (Clavert et al., 1981). Hiện chưa rõ liệu hệ thống mạch máu ở mào tinh hoàn người có được kiểm soát theo cách tương tự hay không.
Theo MacMillan (1954), sự dẫn lưu tĩnh mạch từ thân và đuôi mào tinh hoàn kết hợp lại để tạo thành tĩnh mạch bờ mào tinh hoàn (vena marginalis epididymidis) của Haberer. Các tĩnh mạch này dẫn lưu vào đám rối hình dây leo thông qua tĩnh mạch bờ tinh hoàn hoặc qua các tĩnh mạch cơ bìu hoặc tĩnh mạch ống dẫn tinh. Sự dẫn lưu bạch huyết của mào tinh hoàn xảy ra qua hai con đường (Wenzel và Kellermann, 1966). Bạch huyết từ đầu và thân mào tinh hoàn được loại bỏ theo cùng một con đường như đã mô tả cho tinh hoàn. Các mạch này chạy cạnh tĩnh mạch tinh hoàn trong và cuối cùng tận cùng tại các hạch trước động mạch chủ. Các mạch bạch huyết từ đuôi mào tinh hoàn tham gia cùng các mạch dẫn lưu ống dẫn tinh và tận cùng tại các hạch chậu ngoài.
Sự phân bố thần kinh
Sự phân bố thần kinh của mào tinh hoàn người có nguồn gốc chủ yếu từ các dây thần kinh tinh hoàn trung gian và dưới, lần lượt xuất phát từ phần trên của đám rối hạ vị và đám rối chậu (Mitchell, 1935). Các ống xuất và các đoạn gần của mào tinh hoàn được phân bố thưa thớt bởi các sợi giao cảm (Baumgarten et al., 1968; Baumgarten và Holstein, 1967). Ở các vùng này, các sợi thần kinh chủ yếu liên kết với các mạch máu. Nhiều sợi thần kinh hơn được quan sát thấy ở vùng giữa thân mào tinh hoàn, và mật độ của chúng tăng dần khi tiến dọc theo mào tinh hoàn, trùng hợp với sự xuất hiện và tăng sinh của các tế bào cơ trơn ở các vùng này (Baumgarten et al., 2001 [Ghi chú dịch thuật: văn bản gốc ghi 1971 tại danh mục trích dẫn hình ảnh nhưng ghi 2001 ở đây]). Sự phân bố của các tế bào co bóp và các dây thần kinh giao cảm bên trong mào tinh hoàn có thể giải thích cho các chuyển động nhu động nhịp nhàng của các ống xuất và các đoạn mào tinh hoàn ban đầu, cũng như hoạt động co bóp không liên tục của đuôi mào tinh và ống dẫn tinh trong quá trình phóng tinh dịch (emission) (Risely, 1963). Những sự co bóp sinh lý này là tối quan trọng đối với sự di chuyển của tinh trùng qua mào tinh hoàn.
Cấu trúc tế bào
Biểu mô mào tinh hoàn
Mô học của mào tinh hoàn người đã được tổng quan bởi Holstein (1969) và Vendrely (1981). Nó bao gồm hai loại tế bào chính: tế bào chính (principal cells) và tế bào đáy (basal cells) (được nhìn thấy ở độ phóng đại siêu cấu trúc thấp trong Hình 64.17). Các tế bào chính thay đổi về chiều cao dọc theo chiều dài của mào tinh hoàn do chiều dài của các lông bất động (stereocilia - là các vi nhung mao, không phải lông chuyển). Nói chung, các lông bất động dài (120 µm) được tìm thấy ở mào tinh hoàn đoạn gần, và các lông bất động nhỏ hơn hoặc ngắn hơn (50 µm) được quan sát thấy ở các vùng xa hơn. Nhân trong các tế bào chính kéo dài và thường có các khe lớn cùng một hoặc hai hạch nhân. Phù hợp với quan điểm cho rằng các tế bào chính thực hiện các tiến trình hấp thu và bài tiết, phần đỉnh tế bào của chúng có vô số các hốc có màng bọc (coated pits), các vi túi ẩm bào (micropinocytotic vesicles), các thể nhiều túi (multivesicular bodies), các túi màng có hình dạng không đều, và một bộ máy Golgi rộng lớn. Vì các đặc điểm tế bào này thay đổi dọc theo chiều dài của mào tinh hoàn, nó gợi ý rằng có khả năng hấp thu và bài tiết khác nhau dọc theo chiều dài của ống dẫn (Vendrely và Dadoune, 1988).
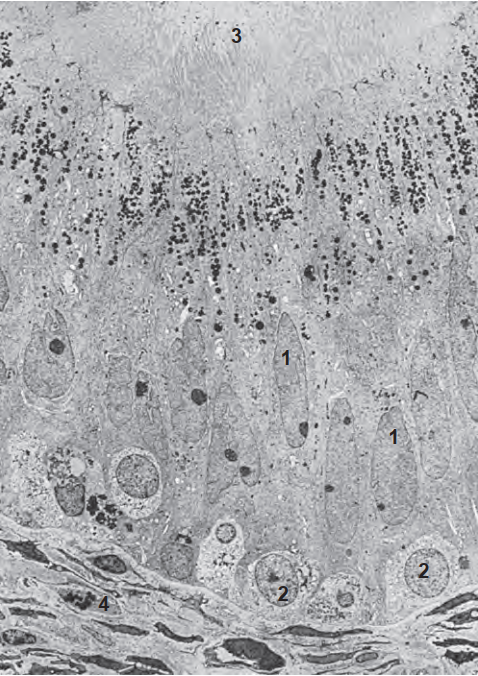
Có ít tế bào đáy hơn nhiều so với tế bào chính lót trong biểu mô mào tinh hoàn. Các tế bào đáy hình giọt nước nằm trên lá đáy và kéo dài về phía lòng ống, phần đỉnh của chúng tạo thành các sợi giữa các tế bào chính liền kề. Chúng được cho là có nguồn gốc từ các đại thực bào và trên thực tế có thể là tiền thân của các tế bào chính. Không giống như các tế bào chính, hình thái của các tế bào đáy duy trì tương đối không đổi trong suốt ống mào tinh.
Biểu mô của mào tinh hoàn thể hiện những sự khác biệt theo vùng dọc theo chiều dài của nó. Bên trong ống mào tinh hoàn chính thức, biểu mô thuộc loại trụ giả tầng và bao gồm các tế bào chính và tế bào đáy như đã mô tả ở trên. Ở đoạn gần, tại vị trí nối của lưới tinh hoàn và các ống xuất, có một sự chuyển đổi rõ rệt từ biểu mô vuông thấp sang biểu mô vuông cao. Biểu mô trong các ống xuất bao gồm các tế bào có lông chuyển và không có lông chuyển (Holstein, 1969). Các tế bào có lông chuyển dẫn tinh trùng từ các ống xuất vào mào tinh hoàn. Các tế bào không có lông chuyển với phần đỉnh nhô ra có khả năng mang bản chất bài tiết và chiếm ưu thế ở các ống xuất đoạn gần (Vendrely, 1981). Các tế bào không có lông chuyển khác có các vi nhung mao gợi ý hoạt động tái hấp thu và chiếm ưu thế ở các ống xuất đoạn xa. Các tế bào không có lông chuyển và có lông chuyển được nối với nhau ở phần đỉnh thông qua các phức hợp liên kết. Điều này gợi ý sự tồn tại của một hàng rào máu - mào tinh hoàn tương tự như hàng rào máu - tinh hoàn (Hoffer và Hinton, 1984; Suzuki và Nagano, 1978). Mặc dù không dày đặc như hàng rào máu - tinh hoàn, hàng rào máu - mào tinh hoàn kéo dài từ đầu đến đuôi mào tinh và có thể đóng một vai trò quan trọng trong việc ảnh hưởng đến thành phần của dịch bên trong lòng mào tinh hoàn ở các đoạn khác nhau của nó (Turner, 1979).
Mô co bóp mào tinh hoàn. Phía ngoại vi của lá đáy của các ống xuất là các tế bào co bóp khác nhau (Baumgarten et al., 1971) (xem Hình 64.17). Ở các ống xuất (vùng xa của đầu mào tinh và vùng gần của thân mào tinh), các tế bào co bóp tạo thành một lớp lỏng lẻo, sâu từ hai đến bốn tế bào, bao quanh ống. Các tế bào này chứa các vi sợi cơ và được kết nối bởi vô số các liên kết kiểu nexus. Ở thân mào tinh hoàn, có các tế bào co bóp lớn hơn với ít liên kết nội bào kiểu nexus hơn, trông giống như các tế bào cơ trơn. Ở đuôi mào tinh hoàn, các tế bào co bóp mỏng được thay thế bằng các tế bào cơ trơn dày tạo thành ba lớp — hai lớp ngoài định hướng theo chiều dọc và lớp trung tâm định hướng theo đường tròn. Lớp co bóp đoạn xa này tăng dần độ dày khi nó hình thành nên ống dẫn tinh. Mô co bóp trong suốt mào tinh hoàn có khả năng tham gia vào quá trình vận chuyển tinh trùng.
Chức năng mào tinh hoàn
Các biến đổi được mô tả trong giải phẫu và mô học của ống mào tinh hoàn từ vùng đầu đến vùng đuôi gợi ý rằng mào tinh hoàn trên thực tế bao gồm vài mô chức năng khác nhau (Vendrely, 1981). Rõ ràng là sự vận chuyển và lưu trữ tinh trùng, khả năng thụ tinh, và sự trưởng thành về khả năng di động là một vài hệ quả của việc đi qua mào tinh hoàn. Điều này được đề cập đầy đủ hơn trong các bài tổng quan của Robaire và Hermo (1988) cùng với Moore và Smith (1988).
Vận chuyển Tinh trùng
Sự vận chuyển tinh trùng qua mào tinh hoàn người đã được tính toán là mất từ 2 đến 12 ngày (Johnson và Varner, 1988). Thời gian di chuyển của tinh trùng qua đoạn đầu - thân mào tinh hoàn nhìn chung tương tự như thời gian di chuyển qua đuôi mào tinh và có khả năng liên quan đến sự sản xuất tinh trùng hàng ngày của tinh hoàn hơn là tuổi của người đàn ông hoặc tần suất phóng tinh (Amann, 1981; Johnson và Varner, 1988). Trong một nghiên cứu, thời gian di chuyển của tinh trùng qua mào tinh hoàn trung bình là 2 ngày ở nam giới có tốc độ sản xuất tinh trùng hàng ngày cao, so với 6 ngày ở nam giới có sản xuất tinh trùng hàng ngày thấp (Johnson và Varner, 1988). Mặc dù tần suất hoạt động tình dục không ảnh hưởng đến thời gian di chuyển của tinh trùng qua đầu và thân mào tinh hoàn, nhưng các "đợt phóng tinh gần đây" có thể làm giảm thời gian di chuyển qua đuôi mào tinh hoàn đi 68% (Amann, 1981).
Vì tinh trùng tinh hoàn bình thường ở người không di động khi chúng đi vào mào tinh hoàn và duy trì tình trạng tương đối bất động bên trong phần đầu mào tinh, các cơ chế khác ngoài sự di động của tinh trùng phải tồn tại để vận chuyển tinh trùng qua mào tinh hoàn. Các nghiên cứu trên động vật đã làm sáng tỏ rất nhiều về khía cạnh này (Bedford, 1975; Courot, 1981; Hamilton, 1977; Jaakkola, 1983; Jaakkola và Talo, 1982). Ban đầu, tinh trùng được mang vào các ống xuất bởi dịch lưới tinh hoàn, và dòng dịch được tạo điều kiện thuận lợi nhờ sự tái hấp thu dịch bởi các tế bào biểu mô ống được gián tiếp bởi thụ thể estrogen. Các lông chuyển có tính di động và sự co bóp của tế bào dạng cơ bên trong các ống xuất cũng hỗ trợ sự di chuyển của tinh trùng. Bên trong mào tinh hoàn chính thức, cơ chế chính chịu trách nhiệm vận chuyển tinh trùng có khả năng là sự co bóp nhịp nhàng, tự phát của các tế bào co bóp bao quanh ống mào tinh.
Lưu trữ Tinh trùng
Sau khi di cư qua đầu và thân mào tinh hoàn, tinh trùng được giữ lại ở đuôi mào tinh hoàn trong những khoảng thời gian khác nhau, tùy thuộc vào tần suất hoạt động tình dục. Ở nam giới từ 21 đến 55 tuổi, có trung bình từ 155 đến 209 triệu tinh trùng hiện diện trong mỗi mào tinh hoàn (Amann, 1981; Johnson và Varner, 1988), và khoảng một nửa được lưu trữ ở vùng đuôi.
Các tinh trùng được lưu trữ ở đuôi mào tinh hoàn, không giống như tinh trùng tinh hoàn, có khả năng di động tiến tới và có thể thụ tinh cho trứng. Khoảng thời gian chính xác mà tinh trùng có thể duy trì khả năng sinh sản bên trong mào tinh hoàn là chưa rõ ràng, nhưng các nghiên cứu trên động vật đã chỉ ra rằng tinh trùng có thể duy trì khả năng sống trong vài tuần sau khi thắt ống dẫn tinh (Hammond và Asdell, 1926; Young, 1929). Tuy nhiên, cũng rõ ràng là khả năng sinh sản của tinh trùng được đo lường in vivo sẽ giảm đi khi tinh trùng được duy trì trong mào tinh hoàn trong những khoảng thời gian kéo dài (Cooper và Orgebin-Crist, 1977; Cuasnicu và Bedford, 1989). Ở người, sự lão hóa của tinh trùng do thời gian di chuyển qua mào tinh hoàn kéo dài và lưu trữ lâu ngày có thể góp phần làm giảm khả năng sinh sản (Johnson và Varner, 1988).
Số phận chính xác của những tinh trùng mào tinh hoàn không được phóng ra là không được biết rõ. Ở động vật, tinh trùng bị mất đi qua sự xuất tinh tự phát, qua việc tự làm sạch bằng miệng (Martan, 1969), trong nước tiểu (Lino et al., 1967), hoặc do sự tái hấp thu của mào tinh hoàn (Amann và Almquist, 1961). Quá trình thực bào tinh trùng bởi các đại thực bào (spermiophages) bên trong lòng mào tinh hoàn đã được quan sát thấy ở người sau khi thắt ống dẫn tinh (Alexander, 1972). Tuy nhiên, liệu cơ chế này có thể loại bỏ số lượng lớn tinh trùng khỏi mào tinh hoàn của những người đàn ông chưa thắt ống dẫn tinh hay không thì vẫn chưa rõ ràng.
Sự trưởng thành của Tinh trùng
Sự di động của Tinh trùng. Tinh trùng đạt được năng lực di động cùng với sự di cư qua mào tinh hoàn. Điều này được quan sát thấy như một sự thay đổi trong kiểu di động và như một sự gia tăng tỷ lệ tinh trùng thể hiện các kiểu di động "trưởng thành". Bedford et al. (1973) lần đầu tiên quan sát thấy hầu hết tinh trùng từ các ống xuất, khi được đặt trong môi trường nuôi cấy, đều bất động hoặc chỉ cho thấy chuyển động co giật yếu ớt hoặc các chuyển động đuôi "chưa trưởng thành" được đặc trưng bởi các nhát đập "quất mạnh" trong các cung rộng dẫn đến rất ít sự tiến về phía trước. Bên trong đoạn mào tinh hoàn ban đầu, kiểu di động chưa trưởng thành này vẫn tiếp diễn. Tuy nhiên, ở vùng thân mào tinh, có sự gia tăng tỷ lệ tinh trùng có kiểu di động "trưởng thành" được đặc trưng bởi các nhát đập có tần số cao, biên độ thấp dẫn đến sự di động tiến tới (Hình 64.18). Bên trong đuôi mào tinh hoàn, phần lớn tinh trùng đã có kiểu di động trưởng thành.
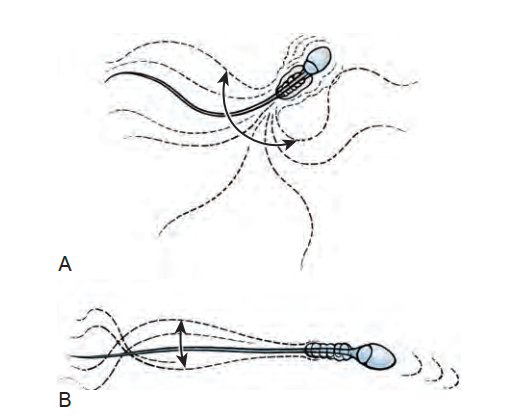
Tầm quan trọng tương đối của thời gian tiếp xúc mào tinh hoàn so với sự trưởng thành theo từng vùng cụ thể đối với việc đạt được các kiểu di động của tinh trùng là chưa được biết rõ. Các nghiên cứu trên động vật chỉ ra rằng sự trưởng thành về khả năng di động, một phần, có thể là một tiến trình nội tại xảy ra độc lập với các tương tác mào tinh hoàn (Horan và Bedford, 1972; Orgebin-Crist, 1969). Các nghiên cứu ở người trên các bệnh nhân bị tắc nghẽn do bất sản ống dẫn tinh bẩm sinh hoặc tắc nghẽn mào tinh hoàn cũng thường xuyên báo cáo khả năng di động kém ở các tinh trùng được hút ra từ mào tinh hoàn đoạn xa và khả năng di động của tinh trùng tốt hơn ở mào tinh hoàn đoạn gần (Matthews et al., 1995; Silber, 1989). Khi kết hợp lại, những quan sát này gợi ý rằng các tinh trùng có thể phát triển khả năng di động dựa trên thời gian tiếp xúc với biểu mô mào tinh hoàn đoạn gần. Tuy nhiên, quá trình trưởng thành này có thể không hoàn toàn giống với quá trình xảy ra thông qua sự tương tác của tinh trùng với mào tinh hoàn trong suốt sự di cư qua tất cả các vùng ống dẫn.
Khả năng sinh sản của Tinh trùng. Tinh trùng tinh hoàn không có khả năng thụ tinh cho trứng trừ khi được tiêm vào trong trứng bằng kỹ thuật vi thao tác (micromanipulation) (Bedford, 1974; Orgebin-Crist, 1969; Yanagimachi, 2005). Ở hầu hết các động vật, khả năng thụ tinh cho trứng của tinh trùng được thiết lập dần dần khi tinh trùng đi qua mào tinh hoàn (Hình 64.19).
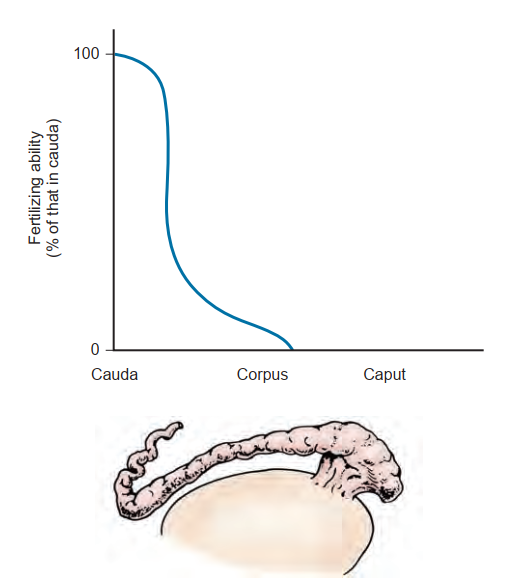
Thật vậy, người ta đã chứng minh ở thỏ rằng tinh trùng từ đầu, thân, và đuôi mào tinh hoàn có thể thụ tinh lần lượt cho 1%, 63%, và 92% số trứng thỏ (Orgebin-Crist, 1969). Các thí nghiệm in vitro ở người sử dụng trứng chuột lang đã bóc màng trong suốt (zona pellucida–free) đã chứng thực những phát hiện này (Moore et al., 1888 [Ghi chú dịch thuật: văn bản gốc ghi 1888, đây có thể là lỗi đánh máy của sách gốc, đúng là 1988]). Trong một nghiên cứu đánh giá năng lực thụ tinh của tinh trùng mào tinh hoàn người, Hinrichsen và Blaquier (1980) đã chứng minh rằng, mặc dù tinh trùng từ mào tinh hoàn đoạn gần có thể liên kết với trứng đã bóc màng trong suốt, nhưng chỉ có tinh trùng từ đuôi mào tinh hoàn mới có thể thực sự xâm nhập vào trứng. Do đó, sự trưởng thành về khả năng sinh sản của tinh trùng, phần lớn, đạt được ở cấp độ thân muộn hoặc đuôi sớm của mào tinh hoàn.
Tuy nhiên, các quan sát lâm sàng gần đây thách thức quan điểm cho rằng sự trưởng thành sinh sản đòi hỏi tinh trùng phải di cư qua toàn bộ mào tinh hoàn. Thật vậy, những bệnh nhân bị tắc nghẽn mào tinh hoàn hoặc bất sản ống dẫn tinh bẩm sinh có thể đạt được thai kỳ tự nhiên sau phẫu thuật nối ống dẫn tinh - mào tinh hoàn (vasoepididymostomy) ở cấp độ các ống xuất (Schoysman và Bedford, 1986; Silber, 1989). Điều này gợi ý rằng tình trạng tắc nghẽn kích ứng sự lệch trục về phía gần của trình tự trưởng thành dọc theo ống mào tinh hoặc có thể có một sự giảm dòng chảy của tinh trùng qua mào tinh hoàn sau các thủ thuật bắc cầu như vậy, cho phép có nhiều thời gian tiếp xúc và trưởng thành tinh trùng hơn (Orgebin-Crist, 1969; Turner và Roddy, 1990). Bất chấp quan điểm này, người ta thường tin rằng khả năng sinh sản sẽ lớn hơn khi miệng nối phẫu thuật được thực hiện ở vị trí càng xa về phía đuôi của mào tinh hoàn (Thomas, 1987). Các phát hiện bổ sung từ việc đảo ngược các ca thắt ống dẫn tinh cũ (>15 năm tắc nghẽn) gợi ý rằng, mặc dù nồng độ tinh trùng phóng tinh sau phẫu thuật được duy trì sau khi nối lại với các khoảng thời gian tắc nghẽn kéo dài, khả năng di động của tinh trùng bị giảm đáng kể. Điều này chỉ ra rằng rối loạn chức năng mào tinh hoàn mắc phải do tắc nghẽn kéo dài có thể đóng một vai trò quan trọng trong tiềm năng sinh sản của nam giới sau phẫu thuật nối ống dẫn tinh (Mui et al., 2014).
Các thay đổi sinh hóa của Tinh trùng. Tinh trùng trải qua nhiều thay đổi sinh hóa khi đi qua mào tinh hoàn (Brooks, 1983). Sự di chuyển qua mào tinh hoàn kích ứng một điện tích màng bề mặt âm ròng (net negative surface membrane charge) (Bedford et al., 1973), và các nhóm sulfhydryl màng tinh trùng bị oxy hóa thành các liên kết disulfide, cải thiện độ cứng cấu trúc của tinh trùng cần thiết cho khả năng di động tiến tới và sự xâm nhập vào trứng (Bedford et al., 1973; Reyes et al., 1976). Các sửa đổi sau tinh hoàn khác của màng tinh trùng bao gồm những thay đổi trong đặc tính liên kết lectin của tinh trùng (Courtens và Fournier-Delpech, 1979; Olson và Danzo, 1981), hàm lượng phospholipid và lipid (Nikolopoulou et al., 1985), thành phần glycoprotein (Brown et al., 1983), độ đáp ứng miễn dịch (Tezón et al., 1985), và các đặc tính iod hóa (Olson và Danzo, 1981). Nhìn chung, những sửa đổi màng này trong quá trình đi qua mào tinh hoàn có thể tăng cường sự gắn kết của tinh trùng vào màng trong suốt (zona pellucida) của trứng (Blobel et al., 1990; Orgebin-Crist và Fournier-Delpech, 1982). Tinh trùng cũng trải qua vô số thay đổi chuyển hóa trong quá trình di chuyển qua mào tinh hoàn (Dacheux và Paquignon, 1980). Chúng bao gồm việc tăng khả năng đường phân (glycolysis) (Hoskins et al., 1975), các thay đổi về pH nội bào và hàm lượng canxi, sửa đổi hoạt tính adenylate cyclase (Casillas et al., 1980), và các biến đổi về hàm lượng phospholipid tế bào cùng các axit béo giống phospholipid (Voglmayr, 1975).
Sự điều hòa chức năng mào tinh hoàn
Các thay đổi của tinh trùng bên trong mào tinh hoàn có khả năng bị ảnh hưởng bởi các dịch và chất tiết bên trong lòng mào tinh hoàn (Chiết xuất từ Blaquier et al., 1989; Robaire và Hermo, 1988). Thành phần sinh hóa của dịch mào tinh hoàn khác biệt so với huyết thanh và cũng thể hiện những sự khác biệt theo vùng về áp suất thẩm thấu, hàm lượng chất điện giải và thành phần protein (Robaire và Hermo, 1988). Những sự khác biệt này có khả năng là hệ quả của các biến đổi trong hệ thống mạch máu, hoạt động của hàng rào máu - mào tinh hoàn, và sự hấp thu cùng bài tiết chọn lọc các chất như glycerylphosphorylcholine (GPC), carnitine, và các axit sialic dọc theo ống mào tinh. Các protein bên trong dịch mào tinh hoàn được biết là có ảnh hưởng sinh lý lên tinh trùng in vitro bao gồm protein di động tiến tới (forward motility protein) (Brandt et al., 1978), yếu tố sống sót của tinh trùng (sperm survival factor) (Morton et al., 1978), yếu tố duy trì di động tiến tới (progressive motility sustaining factor) (Sheth et al., 1981), yếu tố ức chế di động tinh trùng (sperm motility-inhibiting factor) (Turner và Giles, 1982), glycoprotein mào tinh hoàn có tính axit (acidic epididymal glycoprotein) (Pholpramool et al., 1983), và các protein EP2-EP3 kích ứng sự liên kết của tinh trùng vào màng trong suốt (Cuasnicu et al., 1984; Blaquier et al., 1988). Do đó, các biến đổi trong đặc tính dịch ống mào tinh hoàn đóng một vai trò quan trọng đối với sự trưởng thành của tinh trùng trong quá trình di chuyển qua mào tinh hoàn. Do đó, không có gì đáng ngạc nhiên khi mào tinh hoàn là một nguồn tiềm tàng quan trọng gây ra tình trạng rối loạn chức năng tinh trùng và vô sinh nam.
Chức năng mào tinh hoàn được điều hòa bởi hormone. Testosterone và DHT được tìm thấy với nồng độ rất cao bên trong mào tinh hoàn (Leinonen et al., 1980). Điều này gợi ý tầm quan trọng của các androgen đối với chức năng mào tinh hoàn (Brooks và Tiver, 1983). Ở động vật, việc hoạn tinh hoàn dẫn đến không chỉ việc mất các protein mào tinh hoàn phụ thuộc androgen mà còn làm giảm trọng lượng mào tinh hoàn, thay đổi mô học lòng ống, và biến đổi sự tổng hợp cùng bài tiết GPC, carnitine, và axit sialic của dịch mào tinh hoàn. Cuối cùng, mào tinh hoàn bị hoạn sẽ mất khả năng duy trì sự di động của tinh trùng, sự trưởng thành sinh sản và năng lực lưu trữ tinh trùng, các tiến trình này được đảo ngược bằng liệu pháp thay thế androgen.
So với các tuyến sinh dục phụ khác, mào tinh hoàn yêu cầu mức độ androgen tương đối cao hơn để duy trì cấu trúc và chức năng của nó (Prasad và Rajalakshmi, 1976). Các tác động của androgen lên mào tinh hoàn được trung gian chủ yếu thông qua DHT, androgen chính trong mô mào tinh hoàn (Pujol et al., 1976), và/hoặc 5alpha-androstane-3alpha, 17beta-diol (3alpha-diol) (Orgebin-Crist et al., 1975). Thật vậy, điều này được chứng thực bởi thực tế là các enzyme Delta4-5alpha-reductase (xúc tác hình thành DHT từ testosterone) và 3alpha-hydroxysteroid dehydrogenase (chuyển đổi DHT thành 3alpha-diol), vốn sản xuất ra các chất chuyển hóa của testosterone, cũng được tìm thấy trong mào tinh hoàn người (Kinoshita et al., 1980; Larminat et al., 1980). Nó cũng có thể giúp giải thích cho quan sát gần đây rằng việc sử dụng lâm sàng các chất ức chế 5alpha-reductase có liên quan đến việc làm suy giảm chất lượng tinh dịch (Amory et al., 2007).
Chức năng mào tinh hoàn cũng bị ảnh hưởng bởi nhiệt độ (Foldesy và Bedford, 1982; Wong et al., 1982). Việc tiếp xúc mãn tính của mào tinh hoàn với nhiệt độ cao, ví dụ, bằng cách đưa chúng vào bên trong ổ bụng, dẫn đến việc mất các chức năng lưu trữ tinh trùng và vận chuyển chất điện giải. Ảnh hưởng của nhiệt độ lên chức năng mào tinh hoàn có thể giúp giải thích cách thức giãn tĩnh mạch thừng tinh và tinh hoàn ẩn tác động đến vô sinh nam. Các bất thường trong khả năng co bóp của tế bào dạng cơ mào tinh hoàn cũng có thể ảnh hưởng đến chức năng mào tinh hoàn. Ở chuột, việc phẫu thuật cắt bỏ một phần thần kinh của mào tinh hoàn dẫn đến sự tích tụ tinh trùng bất thường bên trong đuôi mào tinh hoàn và làm giảm tốc độ bơi của tinh trùng (Billups et al., 1990). Những phát hiện này có ý nghĩa đối với tình trạng vô sinh do các nguyên nhân bệnh lý thần kinh như chấn thương tủy sống và đái tháo đường.
CÁC ĐIỂM CHÍNH: MÀO TINH HOÀN
- Mào tinh hoàn bao gồm các tế bào chính có chức năng hấp thu và bài tiết, các tế bào đáy có nguồn gốc từ đại thực bào, và các tế bào co bóp giúp tạo điều kiện thuận lợi cho sự vận chuyển tinh trùng.
- Trong quá trình đi qua mào tinh hoàn, tinh trùng trưởng thành bằng cách đạt được khả năng di động tiến tới cùng năng lực liên kết và xâm nhập vào màng trong suốt (zona pellucida) của trứng.
- Chức năng mào tinh hoàn phụ thuộc vào nhiệt độ và androgen (chủ yếu là DHT), đây là những lưu ý quan trọng đối với tình trạng tinh hoàn ẩn, giãn tĩnh mạch thừng tinh, và việc sử dụng chất ức chế 5alpha-reductase.
ỐNG DẪN TINH (VAS DEFERENS)
Cấu trúc đại thể
Ống dẫn tinh là một cơ quan dạng ống có nguồn gốc từ ống trung thận (Wolffian). Ở người, ống dẫn tinh dài từ 30 đến 35 cm, bắt đầu từ đuôi mào tinh hoàn và kết thúc ở ống phóng tinh, nằm ở phía trong so với túi tinh và phía sau so với tuyến tiền liệt. Nó được chia theo kiểu cổ điển thành năm vùng: (1) đoạn mào tinh không có bao xơ nằm trong màng tinh hoàn, (2) đoạn bìu, (3) đoạn bẹn, (4) đoạn sau phúc mạc hoặc đoạn chậu, và (5) bóng ống dẫn tinh (ampulla) (Lich et al., 1978). Trên mặt cắt ngang, ống dẫn tinh bao gồm một lớp vỏ mô liên kết vỏ ngoài chứa các mạch máu và dây thần kinh nhỏ, một lớp áo cơ gồm lớp vòng ở giữa được bao bọc bởi các lớp cơ dọc trong và dọc ngoài, và một lớp niêm mạc bên trong với lớp lót biểu mô (Neaves, 1975). Đường kính ngoài của ống dẫn tinh thay đổi từ 1,5 đến 3 mm, và lòng của ống dẫn tinh không bị tắc nghẽn thay đổi từ 200 đến 700 µm về đường kính (Middleton et al., 2009).
Ống dẫn tinh nhận nguồn cung cấp máu từ động mạch ống dẫn tinh, một nhánh của động mạch bàng quang trên. Sự dẫn lưu tĩnh mạch tương ứng với nguồn cung cấp động mạch. Ống dẫn tinh nhận sự phân bố thần kinh từ hệ thần kinh giao cảm và phó giao cảm (Sjostrand, 1965). Nguồn cung cấp cholinergic dường như không quan trọng đối với hoạt động vận động của ống dẫn tinh (Baumgarten et al., 1975). Có một nguồn cung cấp phong phú các dây thần kinh giao cảm hướng adrenergic bắt nguồn từ dây thần kinh hạ vị chạy qua dây thần kinh trước xương cùng (Batra và Lardner, 1976; McConnell et al., 1982). Các sợi thần kinh adrenergic đã được quan sát thấy ở cả ba lớp của lớp cơ ống dẫn tinh, với mật độ lớn nhất ở lớp dọc bên ngoài (McConnell et al., 1982). Ống dẫn tinh cũng nhận một dây thần kinh adrenergic ngắn (Sjostrand, 1965) và có sự phong phú các thụ thể purinergic trong màng cơ trơn của nó, gợi ý sự đồng truyền dẫn giao cảm và purinergic trong quá trình vận chuyển tinh trùng và phóng tinh (Gur et al., 2007). Các tế bào thần kinh chứa các chất dẫn truyền thần kinh khác, bao gồm neuropeptide Y, enkephalin, galanin, somatostatin, peptide hoạt hóa ruột vận mạch (VIP), và nitric oxide, cũng đã được xác định; tuy nhiên, vai trò của chúng trong chức năng ống dẫn tinh vẫn chưa được biết rõ (Dixon et al., 1998). Đáng chú ý, các mẫu ống dẫn tinh của người thu được khi phẫu thuật nối ống dẫn tinh sau khi thắt ống dẫn tinh cho thấy một sự giảm đáng kể về mật độ của các dây thần kinh hướng noradrenergic của cơ và các dây thần kinh vận mạch bài tiết dưới biểu mô ở các đoạn thuộc tinh hoàn so với các đoạn thuộc bụng. Những thay đổi này có thể ảnh hưởng đến sự vận chuyển tinh trùng tiếp theo trong ống dẫn tinh và do đó ảnh hưởng đến thành công của thủ thuật sau khi phẫu thuật nối lại ống dẫn tinh (Dixon et al., 1998).
Cấu trúc tế bào
Ống dẫn tinh người được lót bởi biểu mô giả tầng (Paniagua et al., 1981). Chiều cao của biểu mô giảm dần dọc theo chiều dài của ống dẫn tinh từ tinh hoàn đến túi tinh. Ngoài ra, các nếp gấp biểu mô dọc đơn giản hơn ở gần tinh hoàn và trở nên phức tạp hơn ở phía xa. Lớp lót biểu mô giả tầng của ống dẫn tinh được cấu tạo từ các tế bào đáy và ba loại tế bào trụ cao, mỏng (Hoffer, 1976; Paniagua et al., 1981). Các tế bào trụ, kéo dài từ nền biểu mô đến lòng ống, bao gồm tế bào chính, tế bào hình bút chì (pencil cells), và tế bào giàu ty thể. Tất cả các tế bào trụ đều thể hiện các lông bất động và nhân cuộn xoắn không đều. Tế bào chính là loại tế bào trụ thường gặp nhất ở ống dẫn tinh đoạn gần, trong khi tế bào hình bút chì và tế bào giàu ty thể tăng dần mật độ ở phía xa. Độ dày của toàn bộ lớp cơ giảm dần dọc theo chiều dài của ống dẫn tinh. Cấu trúc tế bào phức tạp này gợi ý mạnh mẽ rằng ống dẫn tinh không đơn thuần là một ống dẫn thụ động cho sự vận chuyển tinh trùng.
Chức năng ống dẫn tinh
Vận chuyển Tinh trùng. Sự vận chuyển tinh trùng qua ống dẫn tinh bị ảnh hưởng bởi vài tiến trình sinh lý. Đầu tiên, ống dẫn tinh người thể hiện khả năng chuyển động tự phát (Ventura et al., 1973). Nó cũng có khả năng đáp ứng khi bị kéo căng (Bruschini et al., 1977). Cuối cùng, dịch bên trong ống dẫn tinh có thể được đẩy vào trong niệu đạo bằng các đợt co bóp nhu động mạnh mẽ được gợi ra bởi sự kích thích điện của dây thần kinh hạ vị (Bruschini et al., 1977) hoặc bởi các chất dẫn truyền thần kinh hướng adrenergic (Bruschini et al., 1977; Lipshultz et al., 1981). Điều này gợi ý rằng ngay trước khi xuất tinh dịch (emission), với sự kích thích giao cảm, tinh trùng được vận chuyển nhanh chóng từ mào tinh hoàn đoạn xa qua ống dẫn tinh đến ống phóng tinh. Sự vận chuyển nhanh chóng này phù hợp với việc ống dẫn tinh có tỷ lệ cơ trên lòng ống cao nhất (khoảng 10:1) so với bất kỳ tạng rỗng nào trong cơ thể.
Các kho dự trữ tinh trùng trong ống dẫn tinh được ước tính vào khoảng 130 triệu, gợi ý rằng một tỷ lệ đáng kể tinh trùng phóng tinh ở người được lưu trữ trong ống dẫn tinh (Amann và Howards, 1980). Ngoài ra, chất lượng tinh trùng trong ống dẫn tinh, được đánh giá từ những người đàn ông có khả năng sinh sản tại thời điểm thắt ống dẫn tinh, rất giống với chất lượng của tinh dịch phóng ra, với khả năng di động 71% và khả năng sống 91% (Bachtell et al., 1999). Ở thỏ, người ta đã chứng minh rằng trong thời gian nghỉ ngơi tình dục, tinh trùng mào tinh hoàn được vận chuyển qua ống dẫn tinh và rò rỉ vào niệu đạo với số lượng nhỏ (Prins và Zaneveld, 1979, 1980a, 1980b). Điều này gợi ý rằng ống dẫn tinh tham gia vào việc giải thoát mào tinh hoàn khỏi lượng tinh trùng lưu trữ dư thừa. Khi có kích thích tình dục, tinh trùng thỏ được vận chuyển qua ống dẫn tinh tương tự như ở người. Tuy nhiên, sau khi kích thích tình dục, các thành phần trong ống dẫn tinh được đẩy về phía gần hướng tới mào tinh hoàn vì đoạn ống dẫn tinh phía xa co bóp với biên độ, tần số và thời gian lớn hơn so với đoạn phía gần (Prins và Zaneveld, 1980a). Đáng chú ý, với sự nghỉ ngơi tình dục kéo dài, lượng tinh trùng mào tinh hoàn dư thừa một lần nữa được vận chuyển về phía xa, hỗ trợ quan điểm cho rằng ống dẫn tinh quan trọng đối với sự vận chuyển tinh trùng và đối với việc duy trì các kho dự trữ tinh trùng của mào tinh hoàn.
Hấp thu và Bài tiết. Dựa trên cấu trúc tế bào của nó, ống dẫn tinh người có khả năng có các chức năng hấp thu và bài tiết (Hoffer, 1976; Paniagua et al., 1981). Các tế bào chính mang đặc trưng của các tế bào tổng hợp và bài tiết các glycoprotein (Bennett et al., 1974; Gupta et al., 1974). Các lông bất động, hiện tượng tạo bọt ở đỉnh (apical blebbing), cùng các lysosome sơ cấp và thứ cấp bên trong các tế bào chính cũng là đặc tính của các tế bào tham gia vào chức năng hấp thu (Friend và Farquhar, 1967; Murakami et al., 1988). Cuối cùng, hiện tượng thực bào tinh trùng (spermiophagy) bởi các tế bào biểu mô ở bóng ống dẫn tinh đã được quan sát thấy bằng kính hiển vi điện tử quét ở người và khỉ (Murakami et al., 1988). Chức năng ống dẫn tinh bình thường có khả năng phụ thuộc vào androgen vì ống dẫn tinh tích cực chuyển đổi testosterone thành DHT (Dupuy et al., 1979). Việc hoạn tinh hoàn gây ra sự teo — và việc điều trị bằng testosterone gây ra sự phục hồi — cấu trúc tế bào ống dẫn tinh của khỉ (Dinakar et al., 1977), và các sự co bóp tự phát cùng co bóp được kích thích bởi alpha- và beta-adrenergic của ống dẫn tinh chuột bị thay đổi bởi việc hoạn tinh hoàn (Borda et al., 1981). Do đó, mặc dù từng được coi là một ống dẫn cơ trơn đơn thuần cho tinh trùng, ống dẫn tinh hiện nay được nhìn nhận như một cơ quan sinh sản phức tạp.
TÚI TINH VÀ CÁC ỐNG PHÓNG TINH
Cấu trúc đại thể và Cấu trúc tế bào
Túi tinh
Ở người trưởng thành, các túi tinh là một cặp cơ quan tạng rỗng, kéo dài, nằm ở phía sau tuyến tiền liệt và bàng quang. Mỗi túi tinh dài từ 5 đến 7 cm và rộng tới 1,5 cm. Mỗi túi tinh thực chất bao gồm một ống dài 15 cm cuộn xoắn và phức tạp. Ống được cấu tạo bởi ba lớp: lớp lót bên trong là một màng niêm mạc ẩm và có nếp gấp; lớp giữa phần lớn là collagen; và lớp ngoài bao gồm các lớp cơ vòng và cơ dọc chiếm 80% độ dày của thành ống (Nguyen et al., 1996). Niêm mạc của túi tinh, chủ yếu là các tế bào trụ hoặc vuông giả tầng không có lông chuyển, nổi bật với nhiều nếp gấp mỏng, phức tạp tạo ra vô số các hốc (crypts). Ống bài tiết của túi tinh mở vào bóng ống dẫn tinh khi nó đi vào tuyến tiền liệt.
Nguồn cung cấp máu cho túi tinh xuất phát từ động mạch chậu trong và động mạch bàng quang dưới thông qua nhánh tiền liệt - túi tinh (prostatovesicular branch) (Clegg, 1955). Động mạch tiền liệt - túi tinh cũng có thể xuất phát từ động mạch bàng quang trên hoặc từ động mạch thẹn. Thông thường nhất, động mạch tiền liệt - túi tinh có các nhánh trước và sau cung cấp cho các bề mặt tương ứng của túi tinh. Sự dẫn lưu bạch huyết của túi tinh là qua các hạch bạch huyết chậu trong. Các túi tinh được phân bố thần kinh thông qua các dây thần kinh giao cảm từ các dây thần kinh thắt lưng trên và dây thần kinh hạ vị. Sự phân bố thần kinh phó giao cảm xảy ra qua đám rối chậu.
Ống phóng tinh
Các ống phóng tinh là các cấu trúc dạng ống, có bản chất collagen, xếp thành cặp, bắt đầu tại vị trí nối của ống dẫn tinh và túi tinh, chạy qua tuyến tiền liệt, và đổ vào niệu đạo tiền liệt tuyến tại gò niệu đạo (verumontanum). Về mặt mô học, các ống phóng tinh là một sự tiếp nối của túi tinh, ngoại trừ việc lớp cơ vòng bên ngoài không kéo dài vào trong các ống (Nguyen et al., 1996). Có ba vùng giải phẫu riêng biệt đối với ống phóng tinh: phần ngoài tuyến tiền liệt ở đoạn gần; đoạn giữa nằm trong tuyến tiền liệt; và một đoạn ngắn ở phía xa kết hợp với mặt bên của gò niệu đạo trong niệu đạo (Nguyen et al., 1996; Hình 64.20).
Mặc dù ống phóng tinh chứa một lớp cơ bên ngoài ở các đoạn ngoài tuyến tiền liệt và trong tuyến tiền liệt của nó, khi ống chạy về phía xa, lớp cơ bên ngoài tiêu biến, và không có một "cơ thắt" cơ học dạng van nào tại lỗ ống phóng tinh như người ta từng nghĩ trước đây (Nguyen et al., 1996; Hình 64.21). Thay vào đó, sự trào ngược nước tiểu được ngăn chặn và sự tự chủ phóng tinh được duy trì nhờ vào góc nhọn của vị trí cắm của ống vào trong niệu đạo. Lớp biểu mô bên trong của các ống phóng tinh cũng phức tạp và gấp nếp, bao gồm các tế bào trụ đơn tầng và giả tầng. Các ống phóng tinh nhận nguồn cung cấp máu từ các nhánh của động mạch bàng quang dưới và được phân bố thần kinh qua đám rối chậu.
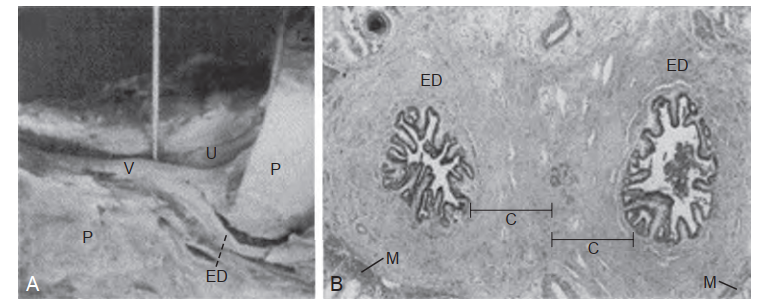
Chức năng của Đơn vị Túi tinh và Ống phóng tinh
Các nghiên cứu trên động vật gợi ý rằng mối quan hệ giữa túi tinh và ống phóng tinh có chức năng tương tự như mối quan hệ giữa bàng quang và niệu đạo (Turek et al., 1998). Túi tinh là một cơ quan cơ trơn có tính co bóp, có độ giãn nở (compliant) với các đặc tính động lực học tương tự như các đặc tính của bàng quang, và ống phóng tinh đóng vai trò như một ống dẫn giống niệu đạo. Lý thuyết này cho phép phân loại tắc nghẽn ống phóng tinh thành hai loại rối loạn, tương tự như tắc nghẽn đường ra của bàng quang: (1) tắc nghẽn do sự tắc nghẽn cơ học của các ống, tương tự như tắc nghẽn đường ra bàng quang và (2) tắc nghẽn "chức năng" của túi tinh, tương tự như rối loạn chức năng tiểu tiện do bệnh lý cơ bàng quang. Ngoài ra, điều này có ý nghĩa đối với việc chẩn đoán các rối loạn ống phóng tinh bởi vì các chẩn đoán hình ảnh giải phẫu "tĩnh", chẳng hạn như siêu âm qua trực tràng, có thể không đủ để phân biệt giữa các rối loạn này, và các thuốc cùng các tình trạng bệnh lý (chẳng hạn như đái tháo đường) có thể làm hệ thống dễ mắc phải tình trạng rối loạn chức năng túi tinh (Smith et al., 2008).
Chức năng túi tinh
Các túi tinh tiết ra một tỷ lệ đáng kể (80%) dịch tinh dịch, và các chất tiết này được tìm thấy trong các phân đoạn sau của tinh dịch phóng ra, sau các chất tiết giàu tinh trùng của mào tinh hoàn và tuyến tiền liệt. Sau khi phóng tinh, tinh trùng đi vào và đi qua dịch nhầy cổ tử cung của nữ giới và tiếp theo là tử cung để đi vào ống dẫn trứng, nơi sự thụ tinh diễn ra. Trong thời gian cư trú ở đường sinh sản nữ, tinh trùng phải trải qua quá trình tạo năng lực (capacitation) trước khi thụ tinh cho noãn. Trong quá trình tạo năng lực, phản ứng cực đầu (acrosome reaction) và sự phát triển của khả năng di động siêu kích hoạt (hyperactivated motility) sẽ diễn ra (Yanagimachi, 1994). Chưa rõ liệu các chất tiết của tuyến tiền liệt hoặc túi tinh có đóng góp vào quá trình tạo năng lực hay không. Trên thực tế, vai trò sinh lý chính xác của dịch túi tinh là chưa rõ ràng, mặc dù ở loài gặm nhấm, nó hoạt động như một nút chặn hoặc hàng rào làm giảm cơ hội để tinh trùng từ một con đực tiếp theo thụ tinh cho noãn. Trước khi phóng tinh, tinh dịch ở dạng lỏng, và sau khi tất cả các thành phần trộn lẫn với các chất tiết của túi tinh, nó sẽ đông vón (coagulates).
Thành phần chính của chất đông vón là semenogelin I, một protein 52-kD được biểu hiện độc quyền ở các túi tinh (Robert et al., 1999). Thông qua việc làm đông vón tinh dịch, các chất tiết của túi tinh có thể thúc đẩy khả năng di động của tinh trùng, tăng cường độ ổn định của chất nhiễm sắc tinh trùng, và đàn áp hoạt động miễn dịch trong đường sinh sản nữ. Chức năng được làm sáng tỏ tốt nhất của tinh dịch người là khả năng cung cấp sự bảo vệ chống oxy hóa cho tinh trùng. Tinh dịch rất giàu các enzyme chống oxy hóa, bao gồm glutathione peroxidase, superoxide dismutase, và catalase (Yeung et al., 1998). Ngoài ra, các phân tử chống oxy hóa taurine, hypotaurine, và tyrosine hiện diện với nồng độ cao (van Overveld et al., 2000). Các hạt lipofuscin từ các tế bào biểu mô đã chết làm cho các chất tiết của túi tinh có màu trắng vàng. Ngoài ra, các chất tiết của túi tinh có tính kiềm và chứa fructose, chất nhầy, vitamin C, các flavin, phosphoryl choline, và các prostaglandin. Nồng độ fructose cao cung cấp năng lượng dinh dưỡng cho tinh trùng. Sự trộn lẫn của chất tiết túi tinh với chất tiết tuyến tiền liệt dẫn đến việc tinh dịch người có độ pH kiềm nhẹ. Tinh dịch có tính axit (pH < 7,2) có liên quan đến tình trạng tắc nghẽn hoặc bất sản túi tinh (Turek, 2005).
CÁC ĐIỂM CHÍNH: ỐNG DẪN TINH, TÚI TINH, VÀ ỐNG PHÓNG TINH
- Ống dẫn tinh có nguồn gốc từ ống Wolffian (trung thận) và vận chuyển tinh trùng từ đuôi mào tinh hoàn đến ống phóng tinh trong quá trình xuất tinh dịch.
- Đơn vị túi tinh và ống phóng tinh tương tự như đơn vị bàng quang và niệu đạo, và có thể bị tắc nghẽn cơ học cùng các rối loạn chức năng dẫn đến vô sinh.
TINH TRÙNG (SPERMATOZOA)
Giải phẫu và Sinh lý học
Tinh trùng người có chiều dài khoảng 60 µm và được chia thành ba phần hình thái: đầu, cổ và đuôi (Hình 64.22).
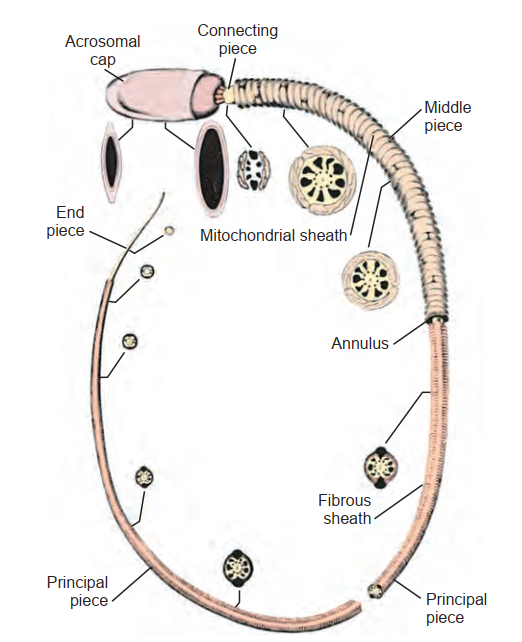
Đầu tinh trùng hình bầu dục, dài khoảng 4,5 µm và rộng 3 µm, chứa một nhân với chất nhiễm sắc được nén chặt cao độ và một cực đầu (acrosome), một bào quan có màng bọc chứa các enzyme cần thiết cho sự xâm nhập qua các lớp vỏ bọc bên ngoài của trứng trước khi thụ tinh (Yanagimachi, 1978). Cổ tinh trùng duy trì sự kết nối giữa đầu và đuôi tinh trùng. Nó bao gồm mảnh kết nối (connecting piece) và trung tử gần. Phức hợp trục sợi (axonemal complex) kéo dài từ trung tử gần xuyên suốt đuôi tinh trùng. Đuôi chứa đoạn giữa (midpiece), đoạn chính (principal piece), và đoạn cuối (endpiece) (Zamboni, 1992). Đoạn giữa dài từ 7 đến 8 µm và là đoạn gần nhất của đuôi, tận cùng tại vòng annulus. Nó chứa trục sợi, với sự sắp xếp vi ống đặc trưng của nó và các sợi dày bên ngoài bao quanh (Hình 64.23).
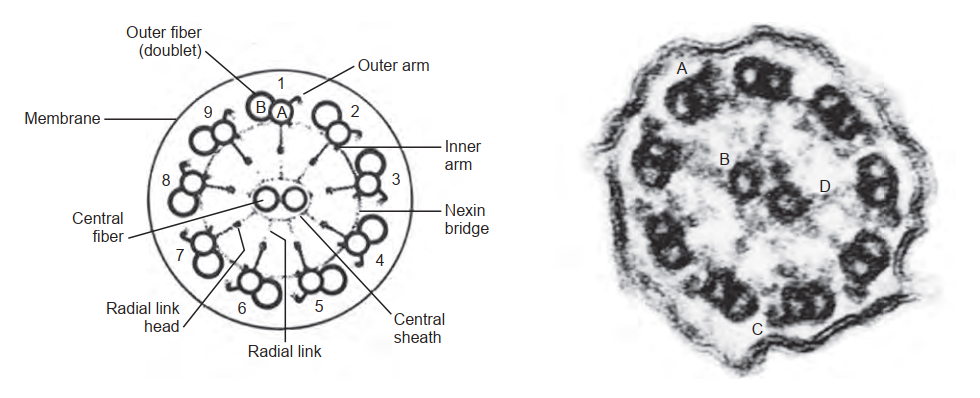
Nó cũng chứa bao ty thể, được sắp xếp theo hình xoắn ốc xung quanh các sợi dày bên ngoài. Các sợi dày bên ngoài, rất giàu các liên kết disulfide, không phải là các protein co bóp nhưng được cho là cung cấp cho đuôi tinh trùng độ cứng đàn hồi cần thiết cho khả năng di động tiến tới (Oko và Clermont, 1990). Tương tự về cấu trúc với đoạn giữa, đoạn chính có vài cột sợi dày bên ngoài được thay thế bằng bao sợi (fibrous sheath). Bao sợi bao gồm các cột dọc và các sườn ngang. Tinh trùng kết thúc ở đoạn cuối, đoạn xa nhất của đuôi tinh trùng, và chứa các cấu trúc trục sợi cùng bao sợi. Ngoại trừ vùng đoạn cuối, tinh trùng được bao bọc bởi một màng tương (plasma membrane) chuyên biệt cao độ có nhiệm vụ điều hòa sự di chuyển xuyên màng của các ion và các phân tử khác (Friend, 1989).
Tinh trùng là một bộ máy chuyển hóa và di truyền phức tạp đến kinh ngạc. 75 ty thể của tinh trùng bao quanh trục sợi chứa các enzyme cần thiết cho quá trình chuyển hóa oxy hóa và sản xuất adenosine triphosphate (ATP), phân tử năng lượng chính cho tế bào. Ty thể là các bào quan sản xuất năng lượng tế bào và cũng có thể gây ra cái chết tế bào theo chương trình (apoptosis) thông qua sự giải phóng cytochrome c. Ty thể được cấu tạo bởi các màng ngoài và màng trong. Màng trong tạo thành các nếp gấp sâu vào trong chất nền, được gọi là các mào (cristae), giúp làm cho diện tích bề mặt của màng trong lớn hơn diện tích bề mặt của màng ngoài. Năm phức hợp chuỗi hô hấp biệt lập nằm trải rộng theo bề ngang của màng trong và là cần thiết cho quá trình phosphoryl hóa oxy hóa: các phức hợp nicotinamide adenine dinucleotide phosphate (NADPH) dehydrogenase, succinate dehydrogenase, cytochrome bc1, cytochrome c oxidase, và ATP synthase. Được chứa bên trong chất nền là các enzyme oxy hóa của chu kỳ axit citric, axit béo, và axit amin; ATP mới được tạo ra; DNA ty thể (mtDNA); và các ribosome.
Ty thể của người chứa DNA (mtDNA) biệt lập với DNA nhân của tinh trùng. mtDNA bao gồm một nhiễm sắc thể vòng, không có histone gồm 16.569 cặp base DNA được sắp xếp thành một sợi nặng duy nhất và một sợi nhẹ duy nhất, và mã hóa cho các protein tiểu đơn vị phức hợp chuỗi hô hấp, các rRNA ty thể, và các tRNA được sử dụng cho sự tổng hợp protein. Các gen này không có các intron. mtDNA cũng dễ bị đột biến hơn nhiều so với DNA nhân (ước tính cao hơn từ 40 đến 100 lần). Các lý do cho điều này có thể bao gồm thực tế là các ty thể nằm gần các phức hợp chuỗi hô hấp và có thể dễ dàng bị tấn công bởi các gốc oxy hóa hoạt động (reactive oxygen species). Ngoài ra, mtDNA không được bao bọc bằng các histone bảo vệ, và các ty thể có cơ chế sửa chữa DNA rất hạn chế (Hirata et al., 2002). Thực tế là mtDNA nhanh chóng tích tụ các đột biến gợi ý về sự cần thiết phải phân hủy tất cả mtDNA của người cha trong trứng đã được thụ tinh. Sự phân hủy này có khả năng được trung gian bởi ubiquitin, một peptide phân cắt protein nhỏ chịu trách nhiệm điều hòa sự phân giải protein ở nhiều mô (Sutovsky et al., 1999).
Từ các nghiên cứu trên động vật, rõ ràng là màng tương bao phủ vùng đầu tinh trùng chứa các protein chuyên biệt tham gia vào sự tương tác giữa tinh trùng và trứng (Saling, 1989). Thật vậy, các protein liên kết carbohydrate trên màng tinh trùng tương tác với protein ZP3 đặc hiệu loài trong màng trong suốt của trứng, dẫn đến trước hết là sự liên kết của tinh trùng vào màng trong suốt và tiếp theo là sự kích ứng phản ứng cực đầu. (Shabanowitz, 1990). Một protein màng tinh trùng khác là PH30, hiện diện trên tinh trùng tinh hoàn, được sửa đổi trong quá trình tinh trùng di cư qua mào tinh hoàn, và hoạt động như một protein hòa màng giữa màng tinh trùng và màng trứng khi thụ tinh (Blobel et al., 1990; Primakoff et al., 1987).
Về mặt sinh lý, trục sợi (axoneme) là bộ máy vận động thực sự và cần từ 200 đến 300 protein để hoạt động bình thường. Trong số này, cấu trúc "9 + 2" của các cặp đôi vi ống (microtubule doublets) ngoại vi và trung tâm là thành phần được hiểu rõ nhất (xem Hình 64.23). Các protein dynein kéo dài từ một cặp đôi vi ống này sang cặp đôi vi ống liền kề và tạo thành các nhánh trong và nhánh ngoài của trục sợi. Trục sợi tinh trùng chứa các enzyme và protein cấu trúc cần thiết cho quá trình chuyển đổi hóa học từ ATP thành chuyển động cơ học và khả năng di động. Dynein là một ATPase kích thước lớn ($2000\text{ kD}$), được kích thích bởi $\text{Mg}^{2+}$, chịu trách nhiệm cho sự trượt vi ống do ATP tạo ra, từ đó gây ra sự uốn cong trục sợi và cuối cùng là chuyển động đuôi của tinh trùng. Cấu trúc dynein có hai hoặc ba đầu chuỗi ngoại vi (chuỗi nặng) hình cầu ($500\text{ kD}$) được nối với một thân chung. Các đầu này kiểm soát chuyển động dọc theo các vi ống. Các nhánh chuỗi trong (chuỗi nhẹ) ($14$ đến $120\text{ kD}$) là các yếu tố hiệu ứng chính của chuyển động và liên kết với các nan hoa xuyên tâm (radial spokes) của bộ máy dynein. Tinh trùng bị đột biến nhánh ngoài sẽ bị giảm khả năng di động, và những tinh trùng bị đột biến nhánh trong sẽ hoàn toàn không có khả năng di động. Các liên kết hoặc nan hoa xuyên tâm kết nối một vi ống của mỗi cặp đôi ngoại vi với cặp đôi trung tâm bên trong và được cấu tạo bởi một phức hợp các protein. Cấu trúc cặp đôi trung tâm bên trong được bao bọc bởi một bao xơ xoắn ốc dạng vòng, nơi các liên kết nan hoa từ các cặp đôi ngoại vi bám vào. Tektins là các protein liên kết với các cặp đôi vi ống ngoại vi, và các liên kết nexin là các protein kết nối các cặp đôi ngoại vi với nhau để duy trì hình trụ của trục sợi.
Kiểu hình cấu trúc tinh trùng khiếm khuyết được công nhận là bệnh lý rối loạn vận động lông chuyển (ciliary dyskinesia). Mặc dù vô sinh là quy luật đối với các trường hợp rối loạn vận động lông chuyển, tinh trùng phóng tinh vẫn có thể di động và nồng độ tinh trùng có thể bình thường. Với kỹ thuật tiêm tinh trùng vào bào tương noãn (ICSI), các trường hợp mang thai lâm sàng và trẻ sinh sống đã được báo cáo sau khi sử dụng tinh trùng bị ảnh hưởng (Cayan et al., 2001). Vì sự di truyền thường là tính trạng lặn nên thế hệ con bình thường là điều có khả năng xảy ra. Nhìn chung, những bệnh nhân bị nghi ngờ có khiếm khuyết cấu trúc tinh trùng sẽ thể hiện khả năng di động của tinh trùng bị suy giảm trầm trọng ($<10\%$). Kính hiển vi điện tử tinh trùng có thể tiết lộ các bất thường về siêu cấu trúc hoặc chức năng của tinh trùng. Các bất thường về cấu trúc tinh trùng hiện được Chemes (2000) phân loại như sau:
- Các bất thường đuôi không đặc hiệu (Nonspecific flagellar anomalies): Đây là bất thường về đuôi thường gặp nhất gây ra tình trạng khả năng di động cực kỳ thấp, thể hiện một kiểu hình cấu trúc của các biến đổi vi ống không đồng nhất, ngẫu nhiên. Những bất thường này có thể phát sinh từ các rối loạn có thể khắc phục được như giãn tĩnh mạch thừng tinh, các gốc oxy hóa hoạt động (ROS), và sự tiếp xúc với các chất độc cho tuyến sinh dục (gonadotoxin). Không có bằng chứng về việc xảy ra có tính chất gia đình.
- Loạn sản bao sợi (Dysplasia of the fibrous sheath): Tình trạng này là một bất thường tinh trùng có tính hệ thống, thường liên quan đến tình trạng bất động gần như hoàn toàn hoặc toàn bộ. Nó có một kiểu hình đồng nhất và đặc trưng hơn, được biểu hiện bằng các biến dạng bao sợi, trục sợi và quanh trục sợi của tinh trùng. Một nhóm nhỏ các bệnh nhân này thể hiện bệnh lý rối loạn vận động lông chuyển điển hình (trước đây gọi là hội chứng lông chuyển bất động), trong đó tình trạng tinh trùng bất động có liên quan đến bệnh lý hô hấp và tật đảo ngược phủ tạng (dextrocardia). Có tỷ lệ xuất hiện cao trong gia đình, gợi ý rằng những tình trạng như vậy thuộc về di truyền.
CÁC ĐIỂM CHÍNH: TINH TRÙNG
- Tinh trùng là các tế bào có lông chuyển, sở hữu cấu trúc trục sợi "9 + 2" cho phép tạo khả năng di động.
- Ước tính có khoảng 200 đến 300 gen điều hòa khả năng di động của tinh trùng.
- Các khiếm khuyết về khả năng di động của tinh trùng, được gọi là rối loạn vận động lông chuyển, rất phổ biến và có thể do nguyên nhân sửa chữa được (bất thường đuôi không đặc hiệu) hoặc do di truyền (loạn sản bao sợi).
- mtDNA tinh trùng người là một vòng DNA tròn, không có histone và không có intron, mã hóa cho các protein phức hợp chuỗi hô hấp và rất dễ bị đột biến.
TÓM TẮT
Quá trình sinh tinh là một tiến trình đặc biệt tinh vi và phức tạp, được thúc đẩy bởi sự bài tiết được điều hòa chính xác của GnRH, LH, và FSH từ trục HPG. Những rối loạn trong môi trường hormone này là nguyên nhân phổ biến gây vô sinh nam. Sự sản xuất tinh trùng ở tinh hoàn hoạt động tối ưu ở nhiệt độ thấp hơn nhiệt độ cơ thể từ 2°C đến 4°C và tạo ra tinh trùng người trưởng thành trong 64 ngày. Các chu kỳ và sóng sinh tinh được tích hợp tốt đảm bảo rằng sự sản xuất tinh trùng ở người duy trì liên tục ở mức khoảng 1200 tinh trùng trong một giây. Sinh tinh là một tiến trình phụ thuộc vào androgen, diễn ra với nồng độ testosterone nội tinh hoàn rất cao. Sản phẩm của quá trình sinh tinh, tức tinh trùng, rời khỏi tinh hoàn dưới dạng các tế bào không di động với năng lực thụ tinh cho noãn bị giới hạn. Sau khi đi qua mào tinh hoàn, tinh trùng thông thường sẽ di động và có khả năng thụ tinh. Trong quá trình phóng tinh, tinh trùng được vận chuyển nhanh chóng từ mào tinh hoàn đoạn xa qua các ống phóng tinh vào trong niệu đạo. Tinh dịch hỗ trợ quá trình chuyển hóa và khả năng di động của tinh trùng, đóng vai trò như một chất chống oxy hóa, và hoạt động như một hàng rào để ngăn chặn các đợt lắng đọng giao tử tiếp theo tiếp cận với trứng.